
9 Postcranial Axial Skeleton
Bill Ryerson
Focus Questions—to Guide Your Reading of This Chapter
- How does the change in rib function over evolutionary time impact the structure of the ribs (and sternum)?
- How can the structure of individual vertebrae tell us about a vertebrate’s locomotion and general body plan?
9.1 Introduction
The axial skeleton is the anchor of the skeleton. It connects the skull and fins/limbs to the rest of the body (with some exceptions). It plays a vital role in the locomotion of vertebrates, serving as the attachment point for many muscles. The axial skeleton houses the spinal cord, protecting it along its travels down the length of the body. Many of the structures of the axial skeleton appear very early in vertebrate history and change greatly across time.
9.2 General Structure and Function
The axial skeleton is composed of the skull, vertebral column, ribs, and sternum (Figure 9.1).
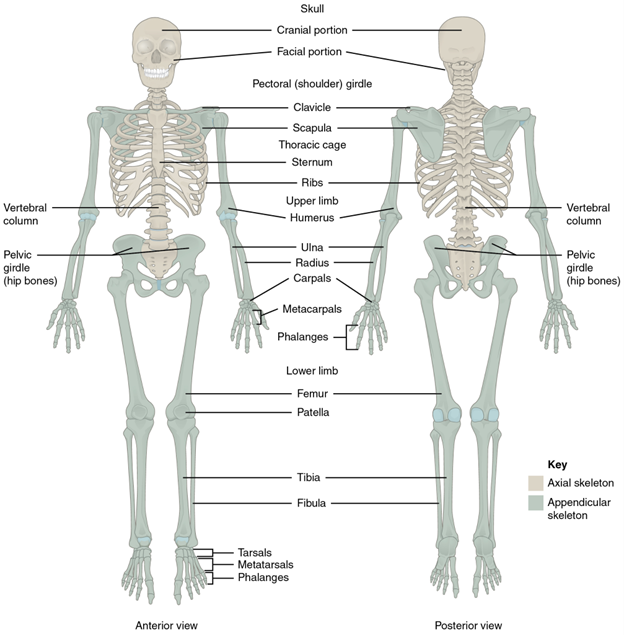
Figure 9.1—The axial and appendicular skeletons of a human.
The skull has already been discussed in Chapter 8, but we shall cover the remaining elements here (what we will refer to as the postcranial axial skeleton). The vertebral column runs from the skull to the end of the tail and attaches to not only the skull but the appendicular skeleton (the fins/limbs and their associated skeletal elements). It is dorsal in the body, typically covered by a few layers of muscle and the integument. The ribs extend from the vertebral column. In groups that have a sternum, it is a ventral structure and the other connecting point for the ribs.
The vertebral column is a segmented structure composed of individual units we refer to as vertebrae. The vertebrae are surprisingly complex structures (Figure 9.2). They provide attachment for epaxial, hypaxial, and other muscles. They have openings for the passages of large blood vessels up and down the body. They also have a single large opening for the passage of the spinal cord. In addition to all these other structures engaging and interacting with the vertebrae, individual vertebrae must also interact and engage with one another. Individual vertebrae moving together at varying degrees drive the side-to-side and dorsal-ventral bending that most vertebrates are capable of (though not all; looking at you, turtles). One of the topics that we will discuss in the evolution of the vertebrae is that of regionalization. Regionalization refers to changes in vertebral shape and number that can characterize a particular region of the vertebral column. The number of regions and the characteristics of those regions are variable between the vertebrate groups and are a useful tool for identifying the vertebrate classes broadly when the limbs/fins or skull are not available. The difference between a lizard skeleton and that of a salamander may not be obvious at first, but there are distinct differences in the vertebral column that can be used to narrow down what it is you are looking at. In fact, vertebrae can be so uniquely shaped that it is possible to identify individual species and estimate their size by the presence of a single vertebra (Box 9.1).
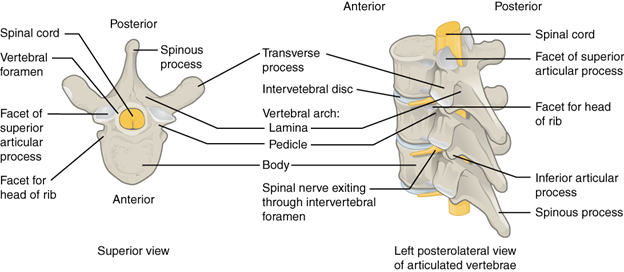
Figure 9.2—Human thoracic vertebrae, illustrating generalized structure and elements of individual vertebrae.
Box 9.1—The World’s Largest Snake…Ever
There are some very large snakes living in the world. The world’s heaviest snake, the green anaconda (Eunectes murinus), can weigh well over 200 lb. The longest snake in the world, the reticulated python (Malayopython reticulatus), can grow to almost 25 ft. in length. Both of these species are dwarfed by the largest snake ever. Known as Titanoboa, this relative of the green anaconda and boa constrictors measures in at over 40 ft. in length, and its weight is estimated at over 2,000 lb. How do we know? Not only do careful measurements of fossil vertebrae from Colombia give us information about its close relatives, but we can also use that information to get an estimate of size. All from one vertebra!
Read more: Giant boid snake from the Palaeocene neotropics reveals hotter past equatorial temperatures.
Between individual vertebrae, intervertebral discs are found. Intervertebral discs are small discs of cartilage with a gel-like center, resisting forces that might cause two vertebrae to come into contact with each other. The intervertebral discs also provide a little bit of space for spinal nerves to pass through canals in the vertebrae to reach the spinal cord.
The ribs are flat in cross section, often curved bones that attach to the vertebrae and the sternum. They are highly group specific in terms of number, shape, and function. Primarily, ribs serve two functions: protecting vital organs (e.g., the heart and lungs in mammals) and anchoring for muscle-directed movement. We will see that what is protected by the ribs and which muscles are attached to the ribs will vary greatly across groups. Some muscles may be more heavily involved in locomotion, while others may be almost entirely respiratory.
The sternum is one part of the axial skeleton that is not consistently apparent in all vertebrate groups. It connects the two halves of the ribs together, forming a “cage” in the tetrapods. It is also assumed to play a supportive and protective role in the tetrapods. The sternum seems to help resist the forces of gravity and body weight when a tetrapod is resting on the ground. The additional bone supports the ribs and helps direct the forces away from the vital organs. Many large aquatic species (the whales and sharks) have either no sternum or a reduced one. When they are beached, their sternum and ribs cannot oppose this newfound hazard of supporting their body weight.
9.3 Development
The postcranial axial skeleton is completely derived from mesoderm, although the different elements do not appear at the same stages. The notochord is the first to appear, a solid rod of mesoderm. After the formation of the notochord, blocks of mesenchymal cells congregate along the notochord (on each lateral side), originating from the sclerotome blocks, which will eventually become the individual vertebrae. The mesenchymal cells will differentiate into chondroblasts and begin the process of creating hyaline cartilage “molds” that will be replaced by bone via the process of endochondral ossification, with the exception of our hagfishes, lampreys, and cartilaginous fishes. The development of the ribs will be largely dependent on the species. For most vertebrates, the ribs originally develop as outgrowths of the vertebral cartilage molds. If these outgrowths are to become ribs, the cartilage mold will “break” off from the vertebrae, becoming its own independent unit. If it remains attached, we will come to know these structures as transverse processes (Figure 9.2). The sternum (in those taxa that have one) develops as an independent unit of hyaline cartilage (also mesoderm) along the midline of the embryo. The cartilage begins as two separate pieces that split and fuse depending on the species before ossifying in its adult configuration.
9.4 Evolution of the Ribs and Sternum
The presence, shape, number, and function of the ribs and sternum are highly variable across the vertebrate groups. It is always useful to keep the phylogenetic history in mind when examining the anatomy, but in the case of the ribs and sternum, we must frequently refer to the behavior of the animal and how these structures must function in the context of behavior. Their structure will also impact the points of articulation with the vertebral column, and so while this section may seem brief compared to that of the vertebral column, you may find yourself referencing it quite a few times.
Hagfishes, Lampreys, and Cartilaginous Fishes
The early diverging vertebrates, from the cyclostomes through the chondrichthyans, do not have any ribs or a sternum to speak of. We might assume that perhaps the foundations of these pieces, like many structures, show up at some point during development and never fully form. However, this does not seem to be the case. The lack of ribs in these groups means that we will also not see the facets (concave attachment sites for a more rounded bone that they articulate with) on the vertebral column that connect the ribs to the vertebral column, making the vertebrae more uniform in appearance down the length of the body than we may see in other groups.
Osteichthyes
In the Actinopterygii (excluding the tetrapods), there is still no evidence of a sternum. However, ribs are present throughout the entire group. Bony fishes have two sets of ribs, aptly named the dorsal ribs and the ventral ribs. While dorsal and ventral are helpful in determining their location, to understand their function and evolution, we need to be more specific, especially in the context of other structures in the body wall. If we imagine sectioning a fish in half and rotating it so that we see the cut, the placement of the ribs will be more obvious (Figure 9.3).
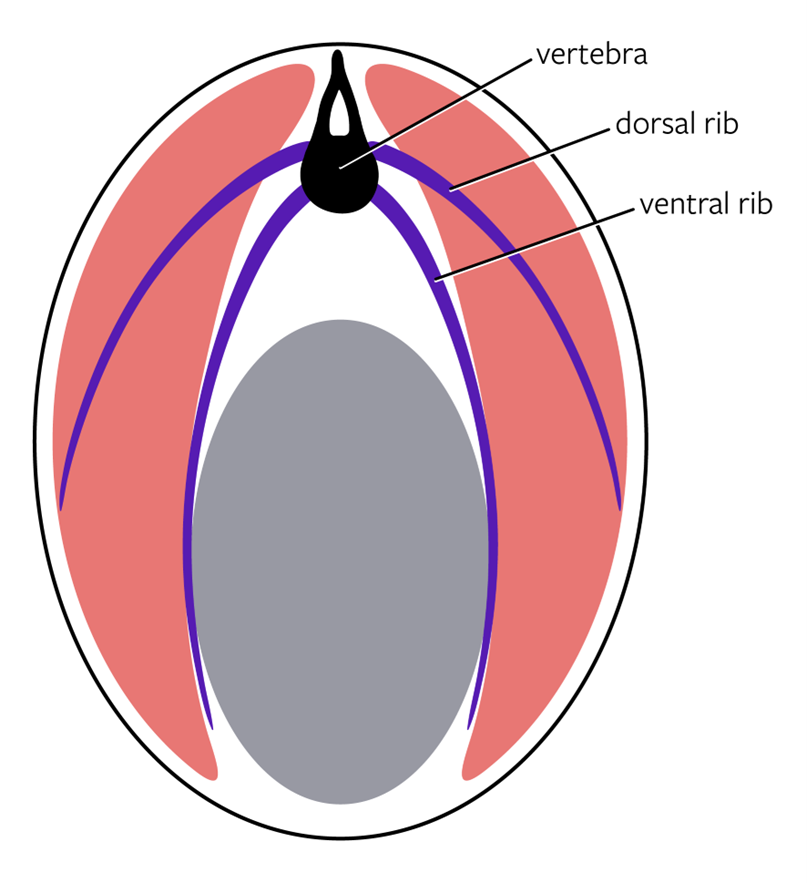
Figure 9.3—Schematic transverse section of a bony fish. The black is the individual vertebrae, with the purple being the dorsal and ventral ribs. The gray is the coelomic cavity, and the pink are axial muscles.
There are several structures apparent to us in this section. At the most dorsal point, we see one of the vertebrae. Around that vertebra, extending downward laterally, are the axial muscles. The axial muscles are named for the placement on the axis of the body (just like the axial skeleton!). Deep to those muscles is the coelomic or body cavity. The body cavity is where all the organs are contained. The two sets of ribs help us delineate these different pieces. The ventral ribs are found between the muscles of the body wall and the body cavity. They serve as an anchor to the body wall muscles as well as protection for the internal organs. The dorsal ribs split the muscles of the body wall (the axial muscles) into two groups: epaxials and hypaxials. We can think of the epaxials as being above the dorsal ribs and the hypaxials as being below the dorsal ribs. There is much more to these muscles than just this slight difference (see Chapter 12 for more), but the differences between the two groups of muscles can be most easily observed by seeing where they are in relation to the dorsal ribs. Without a sternum, the ribs taper off and end either within the axial muscles (dorsal ribs) or with a loose connective tissue connection to the body wall (ventral ribs).
Tetrapods
The transition to land, as you already know, changes many aspects of the skeleton and other systems. The ribs and sternum are no different in this regard, but unlike the other skeletal systems, it can be a little difficult to directly tie changes we see to the functional ramifications of those changes. As of this writing, the origin of the sternum is unclear. We know its current function in tetrapods: Connect the ribs at the ventral ends, reinforce the rib cage, and serve as one element connecting the pectoral girdle to the axial skeleton. These functional demands will drive the variation we see in later groups. When the sternum first shows up in the close ancestors of the tetrapods is still contested. The classic examples of the transitional fossils, Tiktaalik (“fishapod”) and Ichthyostega (stem tetrapod), both lack a sternum. In fact, most fossils that we use to help us understand the evolution of the tetrapods from the rest of the sarcopterygians also lack evidence of a sternum. However, by the time we reach the modern-day amphibians, the sternum is present.
The ribs will also undergo several changes during the tetrapod evolution. The first is that the tetrapods will lose one set of ribs (the dorsal ribs) and the ventral ribs will be modified. The second is that we will begin to see regionalization of the ribs in some groups of tetrapods, with the eventual reduction and loss of a few of these regions by the time we reach the mammals. The modifications to the ribs are highly dependent on locomotion in these groups, so even within these different groups, the ribs will vary greatly in accordance with how that individual species locomotes and what environment it may be found in.
Amphibians
Across the amphibians, the presence and structure of the ribs and sternum are highly variable. The caecilians—the long, legless, and enigmatic amphibians—lack a sternum completely, with long ribs coming off the many vertebrae. Ribs are found all the way from the first vertebrae to the last few vertebrae before the end of the tail. In the frogs, the sternum is highly reduced, serving to connect the two sides of the pectoral girdle together at the center. Most frog species lack ribs as adults, but there is some developmental research that is suggestive of the ribs either fusing to the vertebral column early in development (while still a tadpole even) or vanishing quickly. The salamanders also, unsurprisingly, have considerable variation in their sterna and ribs. Aquatic salamanders, like Necturus, do not have a sternum and have highly reduced ribs. Terrestrial salamanders, like the lungless plethodontid salamanders common to North America, have both a sternum and significant ribs (Figure 9.4).
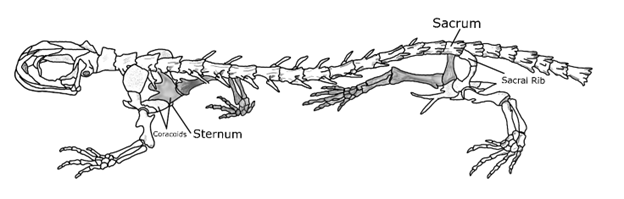
Figure 9.4—The skeleton of the spotted salamander, Ambystoma maculatum.
As was the case with the frogs, the sternum does not connect to the ribs. Instead, it connects the two opposing sides of the appendicular skeleton and in many salamanders is not fully ossified into bone. However, the ribs of the terrestrial salamanders do begin to specialize in ways that will be repeated in other tetrapods. The first is that the ribs are not the same down the entire length of the body (Figure 9.4). Most terrestrial salamanders have ribs that terminate just prior to the pelvic girdle, where the hind limbs attach to the axial skeleton. Moreover, as we get closer to the pelvic girdle, the ribs that are there continually reduce in size. The same is true for the pectoral girdle, as the ribs closest to the forelimbs are also reduced in size. In addition to the changes in size, the ribs also change in their orientation at different segments of the body. Near the pectoral girdle, the ribs “point” ventrally, forming a quasi-cage around the heart and liver. As we move caudally (toward the tail), the ribs themselves also point caudally. The reason for this difference has not been investigated. We assume that it functions in protection of the organs, while the more caudal vertebrae play a different role, perhaps one linked to locomotion. The final item to note about the ribs in salamanders is that we see the emergence of ribs that connect to the vertebrae in two locations. In the bony fishes, both the dorsal and ventral ribs had one connection to the vertebrae, with one head each. In the salamanders, ribs now have two heads, which we can refer to as bicapitate ribs. Each head articulates with a different vertebra.
Nonavian Reptiles
To better understand the evolution of the ribs and sternum, it is necessary to separate the birds from the other reptiles. Flight will greatly impact how the sternum is structured, and there is considerable variation among the other reptiles to consider before we make our way to flight.
The lizards all possess a sternum, with the exception of the legless families (including snakes). The sternum not only connects the appendicular skeleton to the axial but also now serves as an attachment for the ribs (Figure 9.5).

Figure 9.5—The skeleton of a generalized lizard. Note the changes in ribs throughout the trunk region.
For vertebrates where the ribs are connected to the sternum, we can classify them according to how the ribs connect to the sternum. In nearly all cases, there is at least a small branch of hyaline cartilage that runs from the distal end of the rib to the sternum itself. We will refer to this as the costal cartilage. Not every rib has its own costal cartilage. Some ribs will have individual units that merge together to form a single robust costal cartilage that then connects to the sternum. Now we can envision a total of three different categories of ribs, based on their connection to the sternum: one costal cartilage for that rib (true rib), several costal cartilages merging into one (false ribs), or ribs that do not connect to the sternum at all (free/floating ribs). In the lizards with a sternum, we will find examples of all three types of rib. We will also define ribs by their location: cervical ribs for those found anterior to where the pectoral girdle meets the axial skeleton and abdominal ribs for all the floating ribs posterior to the last true/false ribs. The number of ribs that are associated with each region varies considerably. Some lizards do not possess cervical ribs at all, while groups like the monitor lizards may have three or four cervical ribs (Figure 9.6).
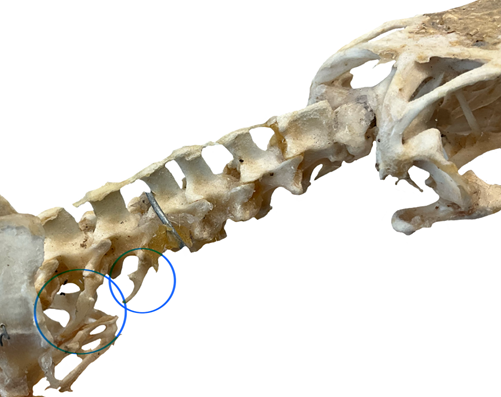
Figure 9.6—The cervical vertebrae of a Savannah monitor (Varanus exanthematicus). Two cervical ribs have been circled in blue.
One of the most interesting changes to occur in the lizards and snakes is not a change in the structure of the ribs themselves but rather one of the functional roles of ribs. To this stage, we have emphasized the role of the ribs in anchoring the axial musculature and introduced their role in protection of the organs. Lizards, having lost the ability to breathe in the same manner that the amphibians have used, must employ different mechanisms for moving air into and out of the lungs (more on this in Chapter 14—Respiratory System). The lizards have employed the ribs for this task. The ribs of lizards can move in several different planes, expanding and contracting the thoracic cavity as needed. Additionally, there is some evidence that rather than moving the ribs as a whole functional unit (as we humans do), some lizards can move individual ribs. Snakes are the undisputed rulers of rib movement. Snakes lack a sternum and the regionalization of the ribs observed in other lizards. Snakes possess ribs along nearly the entire length of the vertebral column, from right behind the head all the way to the cloaca. Ribs in snakes are multifunctional masterpieces. They are integral to protecting the organs, are the only manner by which snakes breathe, and power the many different modes of locomotion that snakes use to get around. However, snakes can also move every rib individually, contributing to their ability not only to move but also to feed. Constricting snakes use their ribs to encircle their prey, generating sufficiently high pressures to kill the prey item. Amazingly, because snakes can move ribs individually, they can constrict their prey with one part of their body and breathe with another!
The turtles, with their robust shells, have moved in seemingly the opposite direction from the snakes and lizards. The ribs and sternum are completely gone in the turtles. What happened to them? Recent fossil and development evidence support the ribs as a key player in the evolution of the shell itself. Several fossil turtles, including Odontochelys and Eunotosaurus, seem to possess large, flattened ribs (Figure 9.7). These flattened ribs are thought to be the precursors to the bony shell of modern-day turtles, with developmental data supporting the transition from individual to the large bony shell.

Figure 9.7—The fossil turtle Eunotosaurus. Two of the ribs are highlighted by white dashed lines.
Leading into the archosaurs, our modern-day alligators, crocodiles, and the extinct dinosaurs, we find a robust sternum with several costal cartilage attachments. The more robust sternum is thought to not only help anchor the larger forelimb bones to the skeleton but also provide additional support for the body when the individual is at rest. Unlike in the lizards, ribs do not move individually in the archosaurs, reducing (but not eliminating) their role in respiration.
Aves
The birds are the next area of considerable variation in the ribs and sternum. The ribs reduce greatly in birds, losing much of their mobility and flexibility in the process. However, the role of the sternum is greatly increased beyond weight bearing and rib attachment. In birds, a key role of the sternum is to serve as an attachment point for the pectoralis muscles. The pectoralis muscles are muscles in the thorax that pull the forelimbs toward the center of the body. For the tetrapod classes we have discussed so far, the pectoral muscles have not been particularly large or of consequence, not impacting the overall structure of the sternum. Birds, being one of the few vertebrate classes capable of powered flight, rely heavily on large pectoral muscles to do so. Large muscles often require large attachment sites, even those that connect via a narrow tendon. In the case of the birds, the large pectoral muscles have had a significant impact on the size and shape of the sternum. The sternum is quite large in most species (Figure 9.8) and can take up more than half the length of the ventral body surface.
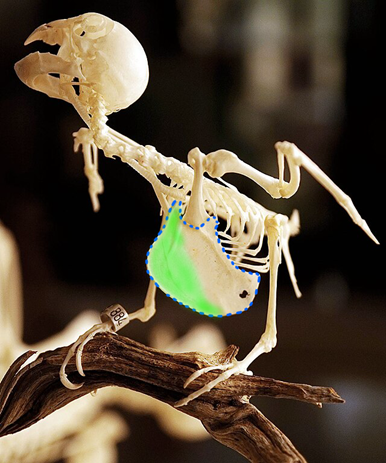
Figure 9.8—The skeleton of a parrot with its large sternum visible, outlined in blue dashed lines. The prominent keel is highlighted in green.
The size of the sternum is more likely a function of stabilizing the ribs and pectoral girdle during flight rather than anchoring the muscles itself. To anchor the muscles, look at the more striking feature of the bird sternum, the large ridge running right down the midline. This is the keel of the sternum and is where the large pectoral muscles attach. The keel can be as large as the rest of the sternum is wide, particularly in birds like hummingbirds and our domestic fowl (e.g., chickens and turkeys). The ostrich, the largest of the flightless birds, has almost no keel to speak of and a reduced sternum relative to its flighted brethren (Figure 9.9).
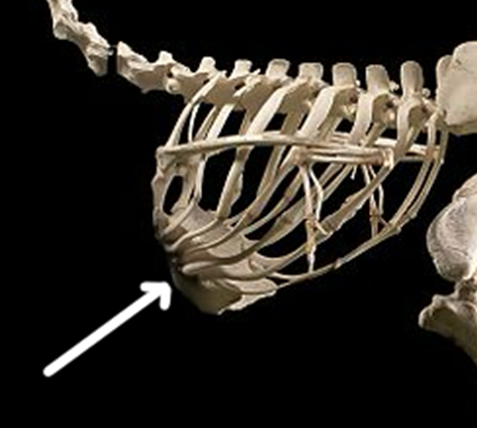
Figure 9.9—The sternum and rib cage of the ostrich (Struthio camelus). Anterior is to the left. The white arrow indicates the sternum.
The ribs of birds are also quite unique. The ribs themselves are a bit broader than in other vertebrates, with very robust connections to the sternum. Bird ribs have uncinate processes, hooked caudal extensions on the ribs. The uncinate processes allow the ribs to hook onto their caudal neighbor. In doing so, the whole rib cage is effectively locked together (Figure 9.10).
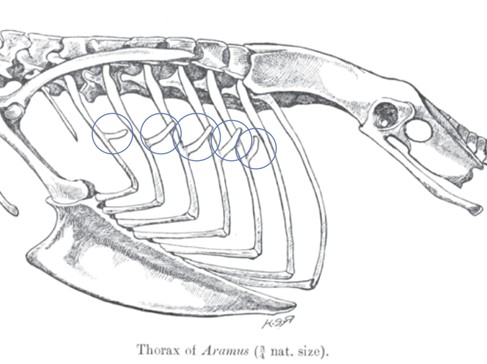
Figure 9.10—The rib cage and sternum of a limpkin (Aramus guarauna). The uncinate processes are circled in blue. Anterior is to the left.
Again, this is an adaptation for flight, as the ostrich (Figure 9.9) seemingly lacks these attachments. This also means that the ribs, being less mobile, play a reduced role in respiration, but birds have developed a highly sophisticated respiratory system that does not rely on the same types of movements (see Chapter 14—Respiratory System for more).
Mammals
The ribs and the sternum of mammals are not strikingly different from those of the other tetrapods but are different in a few key ways (Figure 9.11). The sternum in mammals consists of three individual bones: the manubrium, the body, and the xiphoid. The manubrium is the most anterior/cranial of the three, and it is the attachment point for the clavicle (if there is one) and the first rib. The body is the largest and where the vast majority of the ribs connect. The xiphoid is smaller than the other two components and acts as a connection point for some of the abdominal muscles.
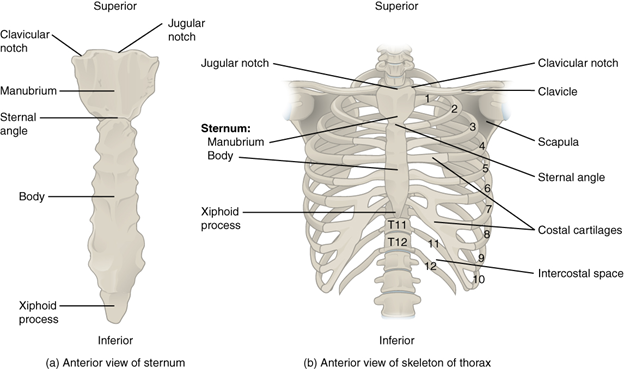
Figure 9.11—The ribs and sternum of the human skeleton.
Mammalian ribs are limited to only about 13 vertebrae in the body with some variation across species. There are no cervical ribs, and the presence of ribs (and their attachment points to the vertebrae) is a distinguishing character that helps separate the different regions of the mammalian vertebral column (more to come).
Mammals possess all three types of ribs: true, false, and floating. The number of each varies by species. The ribs are mobile in mammals, and while individual muscles (the intercostals) have individual units, it appears that no mammals move individual ribs without moving the entire rib cage. The ribs play a role in mammal respiration, but they are not the primary drivers of air movement as we saw in the lizards. The ribs can move to increase the total volume of a single inspiration (take a very deep breath). At rest, they do not contribute to smaller breaths and are not required to move in order to expel extra volume of air.
9.5 Evolution of the Vertebrae
In the evolution of the ribs and sternum, we largely paid attention to how the bones evolved as entire units. As we begin our exploration of the vertebral column itself, we will need to keep in mind not only the column as a whole unit but also how individual vertebrae may change along the way. It is also important to recall the ancestral condition, as represented by the chordate Amphioxus (Figure 9.12).
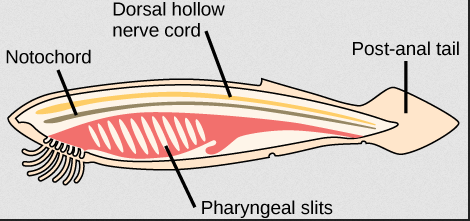
Figure 9.12—Amphioxus, with its prototypical chordate characteristics.
Two of the classic chordate characteristics will help us begin our journey along the history of the vertebral column: the notochord and the dorsal hollow nerve cord. The notochord is a still, rodlike structure composed of cartilage, a single unit running the full length of the body. It is slightly ventral to the dorsal hollow nerve cord (homologous to the spinal cord). With these two elements in mind, we can set the stage for the evolution of the vertebral column.
Cyclostomes
It is admittedly a bit odd to begin discussing the vertebrates with two species that previously have not been considered vertebrates because they seem to lack…vertebrae. The presence of vertebrae in the hagfish and lamprey has been contested for almost two hundred years (literally; see Müller, 1834, Ota et al., 2013, and everything in between). Both groups retain the ancestral notochord, and there are no obvious vertebrae present. However, recent developmental work has found “precursors” to the vertebrae that are found in the caudal region of hagfishes and lampreys. To understand these elements, let us visit Figure 9.13. Looking at this image, use the neural canal (where the spinal cord passes through) as the center of the vertebrae. Ventral to the neural canal is the centrum or body of the vertebrae. Lateral and dorsal to the canal is the neural arch with all its associated processes and facets. We can consider the centrum the “ventral element” of the vertebrae and the neural arch the “dorsal element.” Why are we making this distinction? Because these different elements are what we observe in the hagfish and the lamprey. In the hagfish, along the caudal section of the notochord, several cartilage-based nodules begin to form, homologous to those ventral elements of the vertebrae. In the lamprey, the opposite is true; the dorsal elements are observed just dorsal to the spinal cord itself. However, as they are not fully formed units and do not extend the length of the body, we do not recognize these as vertebrae.
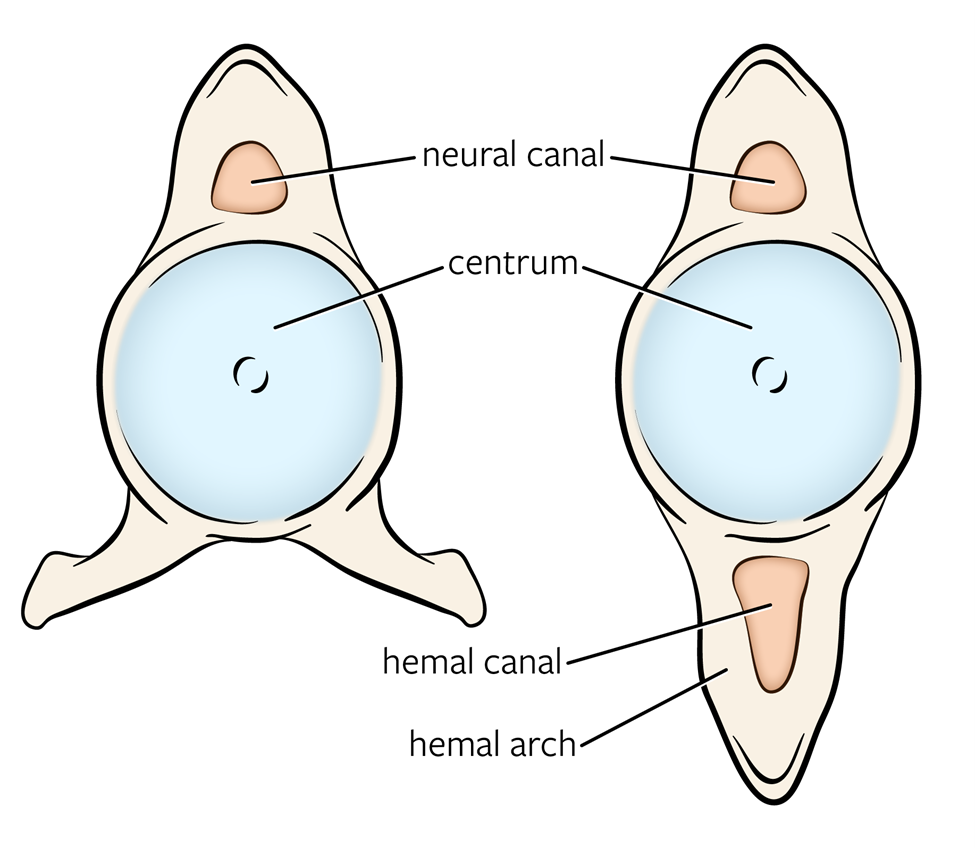
Figure 9.13—The trunk (left) and tail (right) vertebrae of a generalized shark.
Chondrichthyes
The cartilaginous fishes are the earliest group to exhibit true vertebrae, distinct structures from the notochord. They are also the earliest group in which we see at least some regionalization of the vertebral column. Broadly, we can break the vertebral column of chondrichthyans down into trunk and tail vertebrae (Figure 9.13). Both vertebrae have large centra and a neural arch through which passes the spinal cord. There are no attachment points for ribs (articulating facets) because as you recall, ribs are not present in these groups. The shape of the centrum may strike you as a little odd in that it seems to indent (be concave) rather than appear flat like those of humans. This is also a characteristic of many fish, known as amphicoelous vertebrae. These spaces are filled with the intervertebral discs, the remnants of the notochord.
The tail vertebrae seem to have two neural arches, but one of those arches is known as the hemal arch. The hemal arch serves to protect large vessels that run down the tail to feed those large muscles that drive swimming. The hemal arches essentially prevent the tail from bending in such a way that it “kinks” those blood vessels, pinching them off and blocking blood flow. The trunk vertebrae, being less likely to bend to such a degree, do not have the hemal arches. Instead, you may see small processes coming off the ventral surface of the centrum. These processes still border the large vessels but do not completely enclose them. Where both arches are present, they also have large spines (neural spine and hemal spine) that serve as attachment points for the axial muscles mentioned previously. The pectoral and pelvic girdles (where the limbs and fins attach to the axial skeleton) do not attach to the vertebrae directly, and so there are no processes or facets for those attachments. The large sharks show one interesting adaptation to their vertebral column that distinguishes them from the other cartilaginous fishes. Despite being made of cartilage, the vertebrae show a significant amount of ossification, the deposition of mineral salts (calcium, phosphate) into the cartilage. This is different from bone, which would involve different cells, changes to collagen, infiltration of blood vessels, and a few other changes. However, the mineral salts are similar to bone in that they provide strength to the vertebrae, resisting the forces of the large axial muscles. More and more examples of ossified vertebrae in sharks are being found continuously, even in some species that fall outside of the range of “large sharks.”
Osteichthyes
The vertebrae of the bony fishes are very similar to those of the cartilaginous fishes when examined broadly over the entire group. There are trunk vertebrae, and there are tail vertebrae, with the difference between the two types of vertebrae being the same hemal arches as observed previously. In some bony fishes, like the bowfin and the salmon, the dorsal spines can be quite long, much longer than those observed in the sharks and rays. Additionally, the ribs also make an appearance for the first time, and they attach to the vertebrae. The ribs will attach to the vertebrae at locations that we will refer to as articulating facets or parapophyses (hint: wherever you see -pophyses, think vertebrae). These facets are small, smooth surfaces on the vertebrae that allow ribs to move as needed without grinding against the bone of the vertebrae. The parapophyses, depending on the species, may be found on small processes jutting out from the centrum. If you encounter these vertebrae (and ribs), look for a line that separates the parapophysis from the rib. If it appears to be smooth bone throughout the entire process, you have not found the facet yet. The most striking difference found in some fishes is the retention of the notochord. Lungfishes, sturgeon, and coelacanth all retain distinctive elements of the notochord throughout their vertebral column. In these groups, the intervertebral discs are absent; rather, there is a continuous space running through all the vertebrae through which passes the notochord.
Tetrapods: The Transition from Water to Land
As we will observe in many elements of the skeleton, the transition from water to land imposes changes on the structure and function of many bones. For the vertebral column, there is one large change that will impact how the vertebrae are structured and function. In the aquatic environment, a fish (or other vertebrate) has its body weight supported by the density of water. Many fishes can control their buoyancy in the water column and are effectively weightless. Upon moving onto the land, an individual must be able to support its own body weight. Envision a salamander and that long torso between the pairs of limbs. How are they not just dragging their bellies on the ground the entire time? The vertebrae and associated muscles must provide the support needed. Some of these changes may seem minor on their own, but as we move into the different groups, they will impact not only other structures but also how the tetrapods move about on the land. One such structure is called the zygapophyses (also referred to as articular processes). The zygapophyses are predominantly found on the neural arch, often where the neural spine begins (Figure 9.2, articular processes on the human vertebrae). Zygapophyses create points where two vertebrae can link together. Each individual vertebra will have a prezygapophysis (on the anterior side) and a postzygapophysis (posterior). The prezygapophysis from one vertebra articulates with the postzygapophysis of the vertebrae in front of it. This links the entire vertebral column together. While providing stability to the vertebral column, they limit the overall flexibility. The position of the zygapophyses has been used to better understand movement and how different vertebrate groups employ their vertebral column in locomotion.
In addition to the changes across the entire vertebral column, changes to individual vertebrae will also appear. Beyond the increased regionalization of the vertebral column we will observe, two vertebrae will further differentiate to the point that we will give them specific names (Figure 9.14).
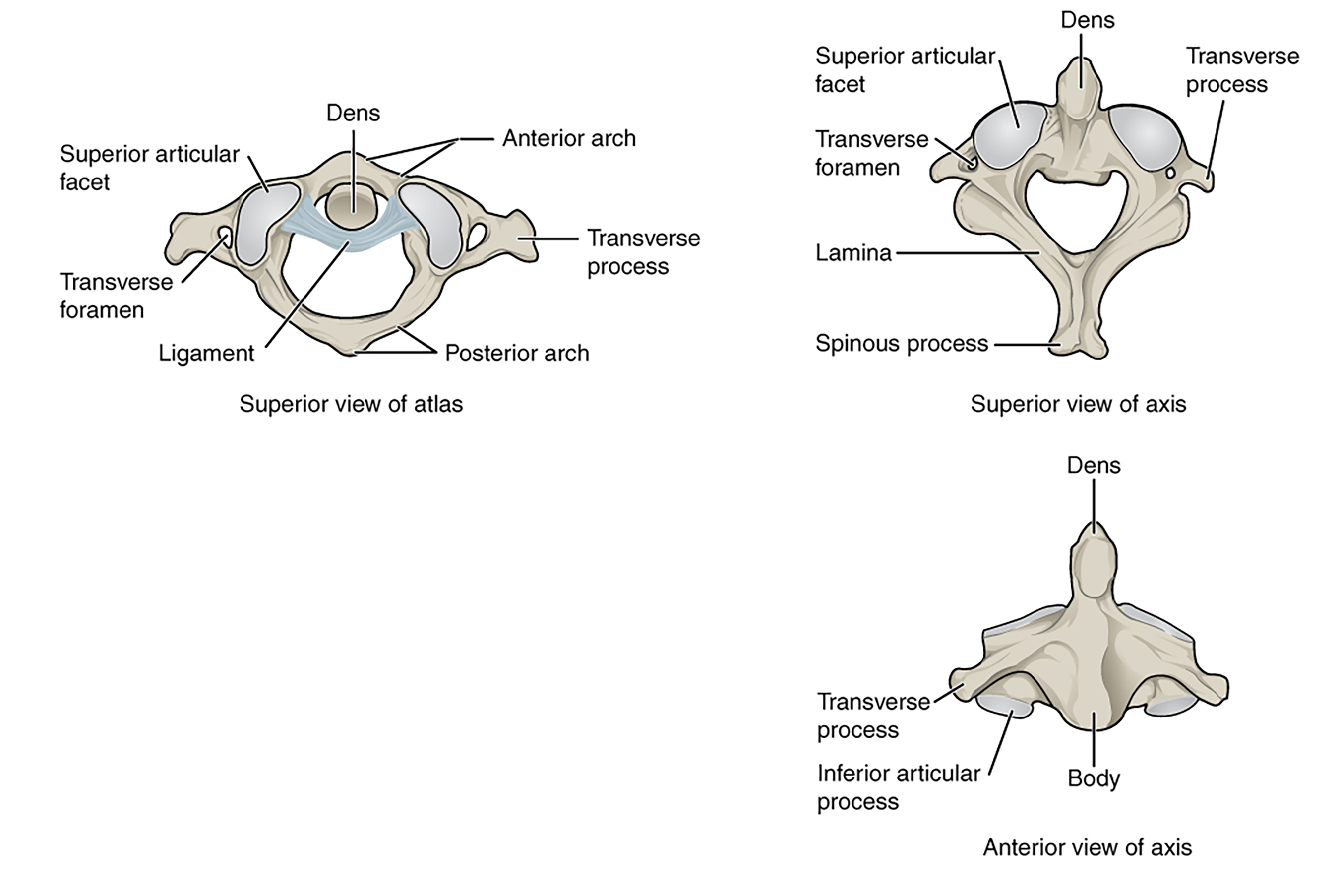
Figure 9.14—The atlas and axis of the human vertebral column. Note that the figure in the top left shows the dens of the axis as it articulates with the atlas.
The first vertebra, the one that connects the skull to the rest of the vertebral column, is given the name atlas. You can remember the atlas by associating it with Atlas, the Greek Titan who held up the sky, as the atlas “holds” the skull. The atlas provides much needed mobility, allowing the skull to move up and down (dorsally and ventrally), a large part of the evolution of the neck (more on this later). In order to pick up food from the ground, movement of the head is needed independent of the rest of the body. The axis, the second vertebra right after the atlas, is also highly modified for the evolution of the neck. The axis has a small process known as the dens or odontoid process that articulates with the atlas. This process allows for rotation of the head. Turn your head side to side, as though you were responding “no” to a question. It is the axis that plays a large role in this rotation in addition to other neck vertebrae. This is where the axis gets its name, by allowing for this rotation. The axis does not appear in every tetrapod group, but those are the exceptions rather than the rule.
Amphibians
With the rise of the amphibians, additional changes to the vertebrae not only distinguish this group as tetrapods but also make them distinct from other tetrapod groups. The first notable change, carried on by other tetrapods, is further regionalization of the vertebral column. In the fishes, we noted the trunk and tail vertebrae, with structural differences between the two corresponding to their function. The amphibians further differentiate regions of the vertebral column: cervical, thoracic/trunk, sacral, and caudal/tail. Cervical vertebrae are those we associate with the neck, running from the skull to the pectoral girdle (the site where the appendicular skeleton connects to the axial skeleton). The number of cervical vertebrae varies in our amphibian groups, one or two in the frogs but can be up to five or six in the salamanders. The atlas is the first cervical vertebra, also referred to as C1. There is no axis in the amphibians even if there is a second cervical vertebra in that species. Some species, like the aquatic mudpuppy salamander, may have ribs that attach to the cervical vertebrae at the articular facets. Between the pectoral girdle and pelvic girdle (where the hindlimb skeleton attaches to the axial skeleton) are the trunk or thoracic vertebrae. The naming of this group is more dependent on the source you are referring to as opposed to convention. In mammals and birds, this will always be referred to as thoracic, while in amphibians and reptiles, it may be referred to as trunk or thoracic. The trunk vertebrae are the sites of attachment for the majority of ribs across all species. Where the pelvic girdle meets the axial skeleton, a single vertebra serves as the connection point. This is known as the sacral vertebra or sacrum (Figure 9.4). No ribs attach to this vertebra, only the bones associated with the pelvis (ilium, ischium, and pubis). The sacral vertebra in salamanders is not highly modified and, with the exception of the ribs, appears similar to the trunk and tail vertebrae. In the absence of articular facets from ribs, small lateral projections are found coming off the centrum. These are known as transverse processes and in many ways function similarly to ribs. They are attachment points for muscles and other bones. The bones of the pelvis attach to the singular sacral vertebra via these transverse processes in both salamanders and frogs.
The salamanders will also see the change in the shape of the vertebrae, at the centrum. In the fishes, we discussed amphicoelous vertebrae, where the centrum of each vertebra was concave on both sides. Many ancestral tetrapods and some salamanders retain this condition. Other salamanders are opisthocoelous: the centrum on the anterior end is convex, and the posterior end is concave. This “tighter” fit to the vertebrae provides stability to the column as the salamander moves about on land.
Frogs possess a unique element to their axial skeleton, shared with no other vertebrate. Caudal to the sacral vertebra, the axial skeleton consists of a single elongated element known as the urostyle. The urostyle possesses a neural arch and canal with the spinal cord passing through it. However, unlike the vertebrae, where many individual units function together, the urostyle is one long piece (Figure 9.15).
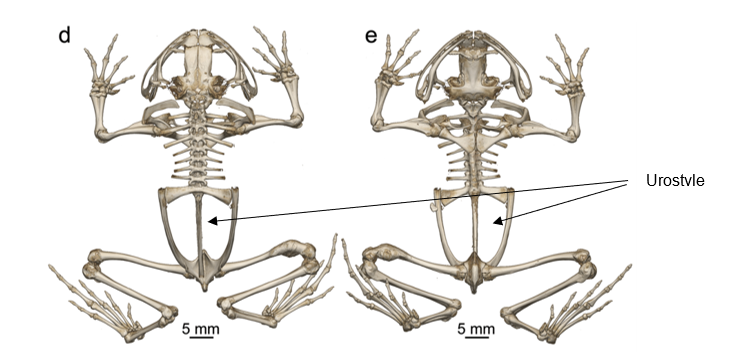
Figure 9.15—The skeleton of a diamond frog (Rhombophryne nilevine). The left image (d) is dorsal and the right (e) is ventral.
This eliminates any mobility in that region for the frogs. The development and evolution of the urostyle remains unclear. As the tadpole metamorphs into the juvenile frog, the caudal vertebrae shrink and fuse together. This is not completely unusual, as it has also been observed in birds and primates. The frog urostyle also has what is known as a hypochord, a developmentally novel structure that enhances the strength and stability of the urostyle.
Nonavian Reptiles
Reptiles have more in common with the salamanders (in terms of their vertebral structure). For the purposes of simplifying our understanding, we can quickly cover the basic plan using the lizards and/or alligators as an example and then move forward to the exceptions (the snakes and turtles). The degree of regionalization is the same with cervical, trunk, sacral, and caudal regions of vertebrae. Ribs do not extend along the entire trunk region, and those without ribs have larger transverse processes than the trunk vertebrae that do. The numbers of sacral vertebrae differ from those in the amphibians, with the reptiles having two sacral vertebrae to the one of amphibians. Alligator caudal vertebrae have hemal arches running almost the entire length of the tail, as do the majority of large lizards. Smaller lizards lack hemal arches. The degree to which caudal vertebrae differ from one another is variable in lizards, correlating with how they use their tail in the environment. Chameleons with prehensile tails, like the charismatic panther chameleon, have regionalization of the vertebrae within the tail itself, an area of active research.
The other difference between reptiles and amphibians is the shape of the centrum. Remember that in fish both the anterior and posterior centra were concave, what we referred to as amphicoelous. Some amphibians retained that shape, while some of the salamanders evolved vertebrae with the opisthocoelous condition, the anterior centrum being convex and the posterior centrum being concave. The reptiles require us to learn a third condition, procoelous. A procoelous vertebra has a concave anterior centrum and convex posterior centrum. Now you should be able to narrow down a lot of skeletons to their vertebrate classes using only a handful of vertebral characters!
Snakes, lacking limbs and therefore girdles (mostly), have lost the regionalization of the vertebral column we discussed in amphibians and other reptiles. Snakes have seemingly “reverted” to a more fishlike pattern, with trunk and caudal vertebrae. There are ribs down the entire length of the body until the cloaca, with articular facets in all the vertebrae along the way. The articular facets for the ribs allow for snake ribs to undertake a large diversity of movements as discussed above.
The turtle vertebral column is also quite the exception. As we discussed with turtle ribs, the development and evolution of the shell have a dramatic impact on the shape and structure of the vertebrae. Turtles have fewer vertebrae than the other reptiles, with the majority of those as cervical vertebrae. The increased number of cervical vertebrae (as we will see in birds) provides increased flexibility. For the turtles, this means an ability to tuck the head into the shell as well as extend it quite far out (Figure 9.16). Trunk vertebrae are fused to the carapace of the shell, at the location of the neural spine (Figure 9.17).

Figure 9.16—The extended neck of the Argentine snake-necked turtle (Hydromedusa tectifera).
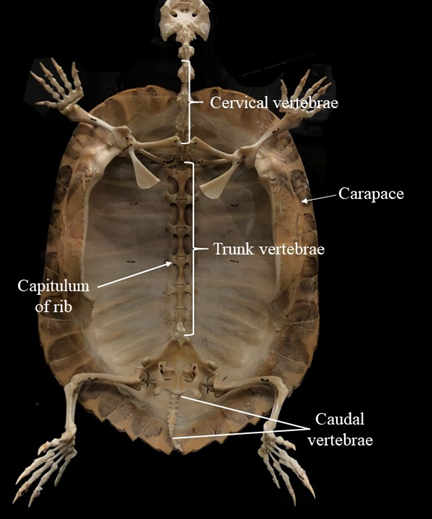
Figure 9.17—The postcranial axial skeleton of a turtle.
Aves
The vertebral column in the birds stands in stark contrast to all the other tetrapods (Figure 9.18). Increasingly, we find greater variation in the vertebrae of the trunk and tail, allowing for a fine balance between supporting the body and allowing for increased mobility. The cervical vertebrae lost their ribs but remained few in number and, with the axis and atlas, did not change their structure significantly. Birds have much more variation in the cervical vertebrae than the other vertebrate groups. Bird cervical vertebrae do not have ribs, with some exceptions. The number of cervical vertebrae found in birds is incredibly variable; 10–26 cervical vertebrae have been counted across bird taxa.
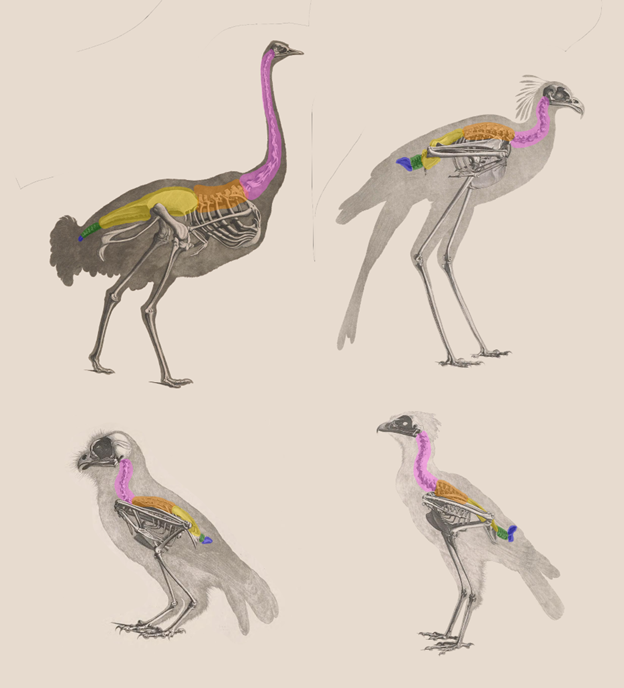
Figure 9.18—Comparative sample of bird skeletons. The regions of the vertebral column are color coded for homologous regions. The birds from top left moving clockwise are an ostrich, a secretary bird, a tropical screech owl, and a crested serpent-eagle.
The large number of cervical vertebrae gives birds incredible flexibility in neck movements, from elaborate courtship maneuvers to the famous ability of owls to turn their heads almost 270 degrees. The incredible variation within bird cervical vertebrae has been suggested to function more like a fifth limb than a region of the vertebral column and may even have subregions that function as independent units. The centra of the cervical vertebrae are also different. We must introduce heterocoelous, a saddle shape to the centra that gives bird necks even more flexibility to maneuver.
For all the variation and flexibility in the cervical vertebrae, the remaining portions of the axial skeleton are fused into distinct units, strengthening the skeleton for the demands of flight. The anterior thoracic vertebrae are fused and articulate with the pectoral girdle into a separate bone called the notarium. The degree and number of fusions are dependent on the group of birds. Interestingly, pterodactyls (not birds but a separate lineage of dinosaurs that evolved flight independently) have converged on a similar arrangement for their anterior thoracic vertebrae and pectoral girdle. This is further support for the role of the fusion of these vertebrae as an adaptation for flight.
The most dramatic alteration of the bird vertebral column occurs further caudally. What appears to be the pelvis of the bird (Figure 9.19) is instead known as the synsacrum.
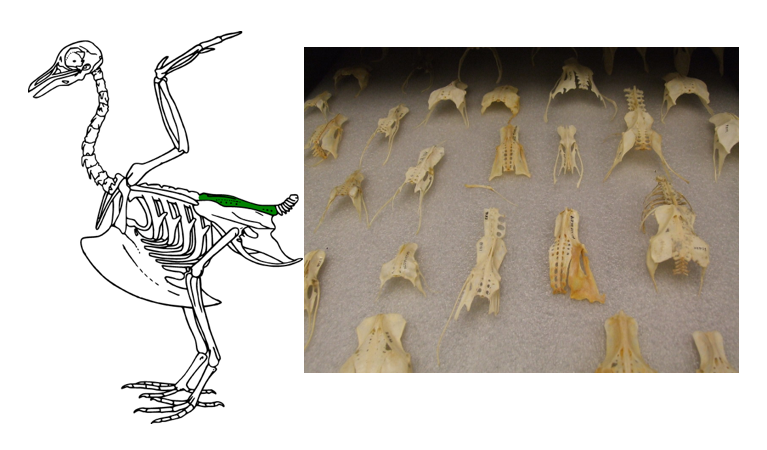
Figure 9.19—Left: The synsacrum (green) on a bird skeleton. Right: Synsacra from multiple bird species.
The synsacrum is an incredible fusion of the lumbar vertebrae (trunk vertebrae without ribs), sacral vertebrae, a few caudal vertebrae, and the three bones of the pelvis. The synsacrum can be quite thin in some bird species, especially the lighter songbirds. The different elements of the vertebral column and the pelvis are still visible, with demarcations more closely resembling the sutures of the skull bones. Posterior to the synsacrum, there are a few caudal vertebrae, some of which may be fused together into a structure that can be called the pygostyle. The pygostyle helps distinguish the skeleton of modern birds from their close extinct ancestors. Archaeopteryx, the most famous ancestor to modern birds, does not have a pygostyle. It has been suggested that the fusion of the vertebrae to make up the pygostyle allows for more independent flight control of the tail feathers, increasing maneuverability in flight.
Mammals
The mammalian vertebral column is highly regionalized, with five recognized regions: cervical, thoracic, lumbar, sacral, and caudal vertebrae (Figure 9.20).
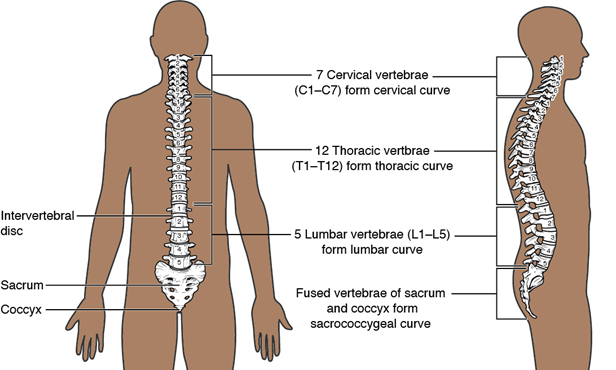
Figure 9.20—The regions of the human vertebral column.
All vertebrae are individually identified by the region they are found in as well as their order in that region with the exception of the caudal vertebrae. For example, the eighth thoracic vertebra is identified as T8. Many of these regions are similar in structure to those observed in the lizards and alligator. With a few notable exceptions, all mammals have seven cervical vertebrae (Figure 9.21).

Figure 9.21—The seven cervical vertebrae of a giraffe (left) and a blue whale (right, circled).
This includes the atlas and axis (C1 and C2, respectively). The thoracic vertebrae are the only vertebrae with ribs and the facets associated with ribs. In mammals, the ribs will attach to two different vertebrae at an articulating facet near the body of two adjacent vertebrae. The lumbar vertebrae are those between the thoracic vertebrae with their ribs and the sacral vertebrae connected to the pelvis. The lumbar vertebrae are characterized by having large transverse processes, attachment sites for many of the axial muscles. The cervical, thoracic, and lumbar vertebrae may also have very large neural spines for muscle attachment, depending entirely on the species. The sacral vertebrae are fused together. Developmentally, they begin as separate vertebrae but are fused together for much of life. There are many openings throughout the sacral vertebrae despite this fusion, allowing for the passage of major blood vessels and nerves. Caudal vertebrae are the most variable across mammals, depending (not surprisingly) on the length of the tail. Even “tailless” mammals such as the primates still retain at least a few caudal vertebrae just posterior to the sacral vertebrae.
One important distinction to make about the mammalian vertebral column involves changes across the vertebrae that correlate directly with changes to the locomotor patterns. To illustrate, let us go back to the salamander and lizard. To emphasize the changes, we will ignore the roles of the limbs in walking on land but instead focus on how the vertebral column moves while the salamander/lizard is moving. In the majority of those animals, they move by bending their bodies side to side. Earlier, we linked this movement to the orientation of the zygapophyses. The zygapophyses were oriented so that their flat portions were parallel to the substrate. This allows the individual vertebrae to glide by one another in the side-to-side motion. However, you can imagine that when any dorsal-ventral motion is required, the zygapophyses will prevent any motion beyond the most minimal. However, in mammals, there is a notable shift in the position of the zygapophyses, especially pronounced in the lumbar vertebrae. The zygapophyses of these vertebrae are oriented laterally, perpendicular to the substrate. Now we have more freedom to move dorsoventrally, at the cost of lateral bending.
This is exactly what we observe. A horse or cheetah at full gallop will illustrate the principle perfectly; note the bending in the lumbar region (Figure 9.22). The bending of the vertebral column dorsi-ventrally helps contribute to the increased stride length in these very fast mammals. In addition, the vertebrae are loosely connected to each other, providing even more flexibility in their movement (Video: BBC—The Cheetah’s Spine).
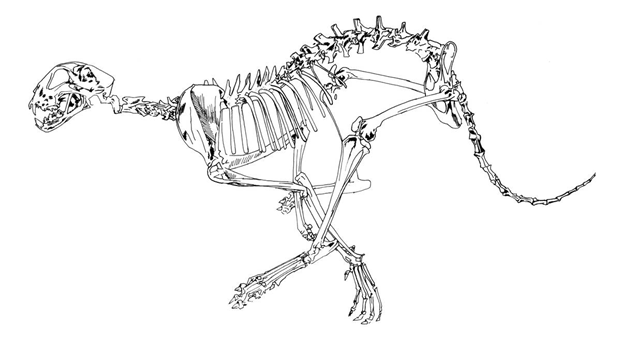
Figure 9.22—An artistic rendering of a cheetah at full gallop, visualizing the dorsoventral bending of the vertebral column.
Humans
As is the case in other mammals, humans have five regions of their vertebral column, with each region having a specific number of vertebrae: 7 cervical, 12 thoracic, 5 lumbar, 5 sacral (fused), and 3–5 caudal vertebrae. The 3–5 caudal vertebrae are highly reduced and fused together, forming a bone known as the coccyx. The vertebral column is curved dorsi-ventrally in humans, with each region curving in opposing directions. The absence of curves or increased curvature in each region is the mark of a pathological condition, named for the region and what is happening to that curve. The vertebral column should not deviate laterally at all; any deviation or curving laterally is identified by the term scoliosis.
Of medical importance to humans is the function and dysfunction of the intervertebral discs. Earlier, we introduced the intervertebral discs as remnants of the notochord, a tough cartilage disc filled with a fluid (nucleus pulposus). Admittedly, after that brief introduction, we essentially ignored the discs until now. The discs act as cushions and spacers between individual vertebrae. This is easiest to demonstrate in humans. With our bipedal posture, our body weight is carried through the legs, hips, and the vertebral column itself. When walking or jumping, the physical forces in the movement press vertebrae together. The discs absorb some of that impact and prevent the vertebrae from coming into contact with one another. Between the vertebrae, pairs of spinal nerves (see Chapter 19 for more) enter and leave the spinal cord. Blood vessels also enter and leave in these spaces. If the vertebrae start knocking into one another, there is a chance that the nerves and blood vessels may become damaged. In addition, the discs themselves are not invincible. They may degrade with time or become damaged by considerable impacts or other forces. There are many colloquial terms that describe disc injury, including slipped disc (technically incorrect), bulging disc, and herniated disc (Figure 9.23).
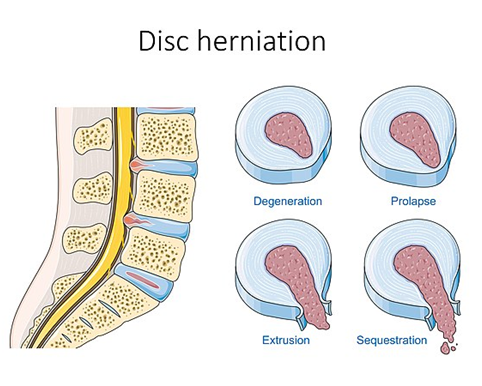
Figure 9.23—Levels of disc herniation in a human lumbar vertebra.
“Slipped disc” is incorrect because the discs grow as part of the vertebral column, and to slip a disc would necessitate tearing at least part of it. All disc damage can be grouped into the category of herniated disc, with differences in severity. A bulging disc tells us that the outer covering is intact, but the nucleus pulposus is pushing outward as the result of increased pressure. A true herniated disc would have the nucleus pulposus leaking out. In the case of a sequestered disc, pieces of the nucleus pulposus have broken off from the rest.
Application Questions
- Why do ribs seem to vanish and shrink in many vertebrate groups?
- How does the orientation of the zygapophyses relate to the locomotion of reptiles and mammals?
- Do changes in the curvature of the human vertebral column at different points result in different medical conditions?
9.6 Further Reading
- Galis, Frietson. “Why do almost all mammals have seven cervical vertebrae? Developmental constraints, Hox genes, and cancer.” Journal of Experimental Zoology 285, no. 1 (1999): 19–26.
- Maddin, Hillary C., Nadine Piekarski, Robert R. Reisz, and James Hanken. “Development and evolution of the tetrapod skull–neck boundary.” Biological Reviews 95, no. 3 (2020): 573–591.
9.7 References
- Aires, Alex S., Leici M. Reichert, Rodrigo T. Müller, and Marco B. Andrade. “Review of morphology, development, and evolution of the notarium in birds.” The Anatomical Record 305, no. 9 (2022): 2079–2098.
- Blanco, M. J., and B. Sanchiz. “Evolutionary mechanisms of rib loss in anurans: A comparative developmental approach.” Journal of Morphology 244, no. 1 (2000): 57–67.
- Brent, Ava E., Emily A. Buchholtz, and Jennifer H. Mansfield. “Evolutionary assembly and disassembly of the mammalian sternum.” Current Biology 33, no. 1 (2023): 197–205.
- Cieri, Robert L. “The axial anatomy of monitor lizards (Varanidae).” Journal of Anatomy 233, no. 5 (2018): 636–643.
- Lowi-Merri, Talia M., Roger BJ Benson, Santiago Claramunt, and David C. Evans. “The relationship between sternum variation and mode of locomotion in birds.” BMC Biology 19 (2021): 1–23.
- Luger, Allison M., Peter J. Watson, Hugo Dutel, Michael J. Fagan, Luc Van Hoorebeke, Anthony Herrel, and Dominique Adriaens. “Regional patterning in tail vertebral form and function in chameleons (Chamaeleo calyptratus).” Integrative and Comparative Biology 61, no. 2 (2021): 455–463.
- Lyson, Tyler R., Gabe S. Bever, Torsten M. Scheyer, Allison Y. Hsiang, and Jacques A. Gauthier. “Evolutionary origin of the turtle shell.” Current Biology 23, no. 12 (2013): 1113–1119.
- Marek, Ryan D., Peter L. Falkingham, Roger BJ Benson, James D. Gardiner, Thomas W. Maddox, and Karl T. Bates. “Evolutionary versatility of the avian neck.” Proceedings of the Royal Society B 288, no. 1946 (2021): 20203150.
- Müller, J. Vergleichende Anatomie der Myxinoiden, der Cyclostomen mit durchbohrtem Gaumen. Erster Theil. Osteologie und Myologie (Gelesen am 4. Und 11. December 1834). Physikalische Abhandlungen der Königlichen Akademie zu Berlin: 65–340.
- Nagashima, Hiroshi, Fumiaki Sugahara, Masaki Takechi, Rolf Ericsson, Yoshie Kawashima-Ohya, Yuichi Narita, and Shigeru Kuratani. “Evolution of the turtle body plan by the folding and creation of new muscle connections.” Science 325, no. 5937 (2009): 193–196.
- Ota, Kinya G., Satoko Fujimoto, Yasuhiro Oisi, and Shigeru Kuratani. “Late development of hagfish vertebral elements.” Journal of Experimental Zoology Part B: Molecular and Developmental Evolution 320, no. 3 (2013): 129–139.
- Scaal, Martin. “Development of the amniote ventrolateral body wall.” Developmental Dynamics 250, no. 1 (2021): 39–59.
- Senevirathne, Gayani, Stephanie Baumgart, Nathaniel Shubin, James Hanken, and Neil H. Shubin. “Ontogeny of the anuran urostyle and the developmental context of evolutionary novelty.” Proceedings of the National Academy of Sciences U.S.A. 117, no. 6 (2020): 3034–3044.
- Zheng, Xiaoting, Xiaoli Wang, Jingmai O’Connor, and Zhonghe Zhou. “Insight into the early evolution of the avian sternum from juvenile enantiornithines.” Nature Communications 3, no. 1 (2012): 1116.
- Secrets of Bones, season 1, episode 2, “Down to Earth,” directed by Sue Doody, featuring Ben Garrod, aired February 25, 2014, on BBC Four, British Broadcasting Company.
