
19 Peripheral Nervous System
Bill Ryerson
Focus Questions—to Guide Your Reading of This Chapter
- The cranial nerves are named for their number, structure, and function in mammals. How might this mislead you when you examine the cranial nerves in other vertebrates?
- The nervous system interacts with many other different systems, most of them covered in other chapters in this book. How are the other systems controlled and/or monitored using the peripheral nervous system?
19.1 Introduction
When most of us are asked to think about the nervous system, our immediate response is to consider the brain. This is not the wrong response but rather a simple fact—the rest of the nervous system does not immediately jump to mind. The brain is selfish, it only considers itself, but that is a question for metaphysics and psychology. We will instead force the brain to consider many of the other elements of the nervous system beyond itself. We consider the spinal cord a part of the central nervous system (CNS), as it shares many developmental and structural similarities to the brain, and rightfully the two are grouped together. The remainder of the nervous system is broadly categorized as the peripheral nervous system (PNS), which includes the spinal nerves and the cranial nerves. With a few notable exceptions, what these different structures all have in common is that they serve as messengers, carrying signals to/from the brain and the other parts of the body. They may carry signals from the senses to the brain or carry signals from the brain to the muscles to create a movement. In doing so, they serve as important links across the body. We can consider these elements part of the rapid-response team of the body as compared to the endocrine system (Chapter 21), which may have longer-lasting impacts. To further this interconnectedness one step forward, the endocrine and nervous systems are not independent units; action of the endocrine system impacts the nervous system and vice versa. In this chapter we will cover the general elements of the PNS, examining the structure and function over evolutionary time. We will pay especially close attention to the cranial nerves, as their story is compelling not only in their function but in how they are incredibly informative in the story of evolution as well as a key diagnostic tool in human and animal health. We will discuss small elements of nervous system physiology here as needed, but know that there are many aspects of physiology that are beyond the scope of this anatomy textbook.
19.2 Generalized Structure and Function
The spinal nerves and the cranial nerves are structurally similar in many ways. They are nerves, with cell bodies, axons, and dendrites (mostly), and interact with other structures at synapses. They carry signals to and from the CNS via discrete pathways. The key difference between the two sets of nerves is their origin. Spinal nerves arise along the spinal cord, between pairs of vertebrae. The majority of cranial nerves connect directly with the brain itself, with either the forebrain or the brain stem, and leave through the skull. There is one exception to this rule, but you will have to contain your suspense for now. So why do we make the distinction between the cranial and spinal nerves? There are two key reasons for this separation, both of which involve the origin and path of the nerves. Cranial nerves, in addition to originating directly in the brain, remain distinct from other nerves from origin to insertion. The spinal nerves, in contrast, merge with other spinal nerves shortly after emerging from their origin on the spinal cord. In some regions of the tetrapod body, these mergers form large distinct structures known as plexuses. We will discuss the plexuses in more detail later, but keep this in mind as we start to explore the different nerves.
Both cranial and spinal nerves can be further subdivided based on their function. Moving forward, we will discuss the difference between where the signals originate and where they go (sensory nerves, motor nerves, and interneurons). We can also classify nerves based on how they contribute to how the body functions. For this, we can group nerves into whether they are part of the “rest-and-digest” or “fight-or-flight” systems.
The first and most direct way to break nerves down is determining in which direction the nerve is headed (Figure 19.1).
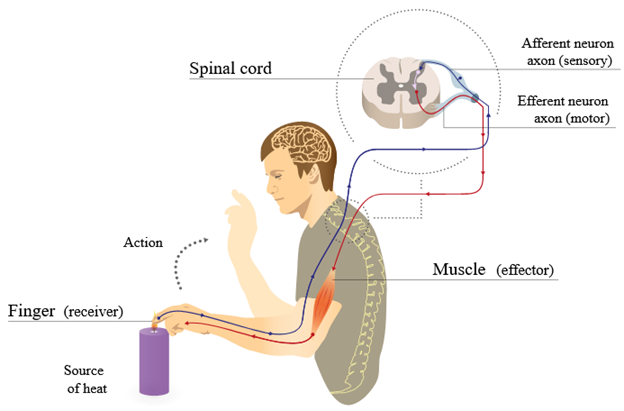
Figure 19.1—The sensory and motor pathways for nerves in one reflex.
Axons can only carry signals in a single direction, so while we may have some difficulty determining direction in a large nerve, we know that individual axons are unidirectional. If an axon is carrying a signal toward the CNS, then we might refer to the whole cell as a sensory or afferent neuron. Signals that are carried away from the CNS do so on neurons that we can refer to as motor or efferent neurons. This is helpful more in a theoretical sense than a practical sense. This helps us visualize how the entire nervous system functions together. Information is carried from a sensory receptor (more on those in Chapter 20) to the CNS, where the integration and processing of signals occur. A response is sent out to an effector, which could be lots of different tissue types. Sensory in, motor out: This is the nervous system in a nutshell.
Now it’s just a matter of looking at where the signals come from, how the signals arrive, what is done with that information, and how the response is carried. This is where the true complexity of the nervous system comes into play. How these all function together is a little beyond the scope of this textbook, but it does help us separate structures out. However, you can still isolate different aspects of these subdivisions to help your understanding of the nervous system as a whole. You can see a great example of this in Figure 19.2.
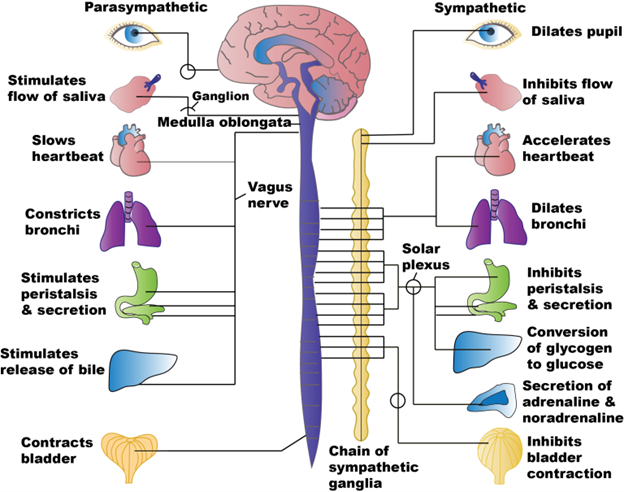
Figure 19.2—The parasympathetic and sympathetic divisions of the autonomic system in humans.
In mammals, we can divide the autonomic nervous system (the components of the nervous system that regulate structures not under voluntary control) into the parasympathetic and sympathetic divisions. The parasympathetic system oversees the regulation of digestion and excretion and slows the heart rate, giving it the colloquial name of the “rest-and-digest” system. Activating the sympathetic division will increase heart rate, slow digestion, and cause the release of adrenaline, hallmarks of the “fight-or-flight” system. Each division has different pathways (and different nerves) that carry their signals. These different nerves may eventually reach the same organs (e.g., the heart), but they have different impacts on the target organ (increasing or decreasing heart rate).
As we just discussed, individual neurons can be defined as afferent or efferent, sensory or motor, but whole nerves are a mix of both types. However, there are a few locations where the entire nerve consists of only sensory or motor nerves. One such location is right where the spinal nerves enter/leave the spinal cord. After passing through the meningeal cuff (an extension of the meninges associated with the spinal cord), nerves split into two pathways depending on whether they are sensory or motor (Figure 19.3).
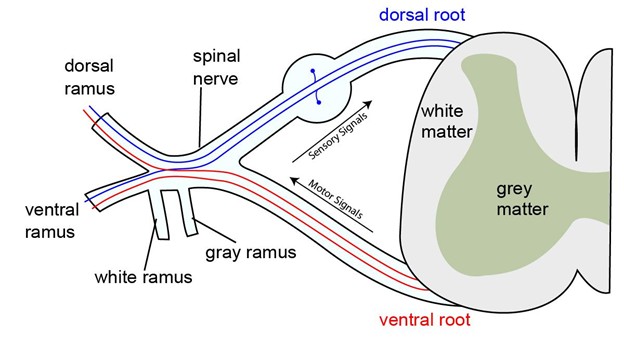
Figure 19.3—The dorsal and ventral roots leaving the spinal cord.
These pathways form what we know as nerve roots and are given the same naming as the horns of the spinal cord from above: dorsal root and ventral root. This is one of the only locations where you can reliably identify a nerve as either sensory or motor in most vertebrates. The dorsal root contains only axons of sensory/afferent neurons, while the ventral root contains only axons of motor/efferent neurons. In this way you can visualize the generalized function of the entire nervous system. Signals from sensory receptors travel through spinal nerves and move through the dorsal root. Entering the dorsal horn, the signals are processed/integrated, and a response is formulated. The response is generated in the ventral horn and leaves through the ventral root. It is then carried via spinal nerves to the target organ/muscle/gland. Note in Figure 19.3 that it seems the dorsal root and ventral root merge together right after leaving the spinal cord, and that is because they do. This condensed version of nervous system function is surprisingly similar to how reflexes work.
“What about the remaining regions of white matter in the spinal cord?!” you shout. They too can be divided into regions as they are divided by the gray matter horns. The different areas of white matter are known as funiculi (Latin for “string,” a reference to the white nerves) or sometimes called columns: dorsal funiculus, lateral funiculus, and ventral funiculus. Their role is more variable across vertebrates, but there are a few generalizations we can make here. Remember that they are considered white matter as they consist of myelinated axons. Therefore, they are carrying action potentials (nerve signals) up, down, and across the spinal cord. Different regions within these funiculi are referred to as tracts, either ascending or descending. Keep this in mind when considering their function. In most groups, they make up pathways for signals coming from or to different parts of a body. There is some new terminology here, so do not hesitate to take your time and consider the wording carefully. One distinction that we have not discussed but is important here is the difference between somatic and visceral. The somatic and visceral components of the PNS represent different types of targets. To simplify the point for now, think of the somatic system as working with the skeletal muscles (voluntary) and the visceral system for all the internal organs (involuntary). Let us combine that with the sensory and motor roots/horns and note that we end up with four different ways to classify nerves in the PNS: somatic sensory, somatic motor, visceral somatic, and visceral motor. And in fact, these are what the different funiculi can be divided into. The dorsal funiculus has somatosensory tracts (ascending to the brain), and the ventral funiculus has a large somatomotor tract (descending). The lateral funiculus contains two tracts: an ascending visceral sensory and descending visceral motor tracts. While this overall pattern is consistent across vertebrates, as we will discuss later, there is some variation here as well.
19.3 Development
In the previous chapter, we glossed over one element of the development of the nervous system that is very key to the development of the spinal and cranial nerves. During neurulation, small groups of cells split off from the folding neural tube. Neural crest cells are a population of mesenchymal cells that originate from the dorsolateral surface of the neural tube, between the dorsal surface of the neural tube and the epidermis just above it. Neural crest cells have multiple functions (Chapter 4), but they play a huge role in the formation of the peripheral nervous system by giving rise to all the cranial and spinal nerves. The neural crest cells will migrate all over the body and are regulated by a whole host of gene and transcription factors that will ultimately determine what they will become. Neural crest cells that migrate in the trunk region will play a large role in the development of the nerves in that region, including the sympathetic and parasympathetic pathways. Several of the ganglia (groups of neuron cell bodies outside of the brain) of cranial nerves are formed by these neural crest cells as well. This is still a very active area of research, with new work appearing monthly.
19.4 Evolution of the Spinal Nerves
It may come as a surprise to hear that the spinal nerves do not greatly change over evolutionary time in the same manner as many of the other structures discussed in this book. There are a few large transitions that occur in the transition from water to land and with the development of paired appendages.
Cyclostomes
There is one key element to the spinal nerves in hagfishes and lampreys that must be addressed. In the previous section, we discussed that in most vertebrates it is extremely difficult to distinguish between sensory and motor nerves, as the two groups will travel together in intermixed nerves. The emphasis there was on most. The hagfish and lamprey are the exception to this rule. The spinal roots containing sensory neurons and motor neurons remain separate as they become spinal nerves. These separate spinal nerves continue to their targets completely separate. This is one of the reasons we understand the nature of the dorsal and ventral roots and horns and how they translate into sensory systems and muscle activity.
Chondrichthyes and Osteichthyes
The major transition in the jawed fishes is that we now see the intermixing of the dorsal and ventral spinal roots when they emerge as spinal nerves. Coinciding with these changes are also changes to the skeletal system that impact the structure of the spinal nerves. Beginning with the cartilaginous fishes, the notochord/vertebral column is no longer a single structure that runs the length of the body. Instead, we have discrete blocks of bones (though recall that they are made of cartilage in Chondrichthyes) known as the vertebral column. Individual blocks are called vertebrae. Between individual vertebrae, there is an intervertebral disc, resulting in a little space between the vertebrae. This, along with notches on the vertebrae themselves, creates space for the spinal nerves to leave the spinal cord. These spaces also allow for individual movement of vertebrae during locomotion that does not impact the nerves themselves.
Tetrapods—Amphibians and Reptiles
When the tetrapods emerge onto land, the factors that influence the structure and function of the spinal nerves result in some significant changes. The evolution of limbs greatly changes the structure of the spinal nerves. The limbs, with their increased range and diversity of movements, required a series of major changes to the skeletal (Chapter 10), muscular (Chapter 11), and now the nervous system.
At the beginning of our discussion, one of the major distinctions between spinal and cranial nerves is that cranial nerves don’t merge with each other, while spinal nerves do. While the spinal nerves do merge closer to the spinal cord in more basal vertebrates, we see more complex patterns of merging in tetrapods. There are several areas of complex merging to be found, but we will focus on the limbs and see that the spinal nerves that innervate them will change drastically. These groups of nerves will form a network that individually we can call a plexus (Latin, “braid”). There are several plexuses found in tetrapods, associated with the limbs. The forelimbs are innervated by the brachial plexus. “Brachial” typically refers to the forelimbs, and this is no exception. Figure 19.4 illustrates the human brachial plexus.
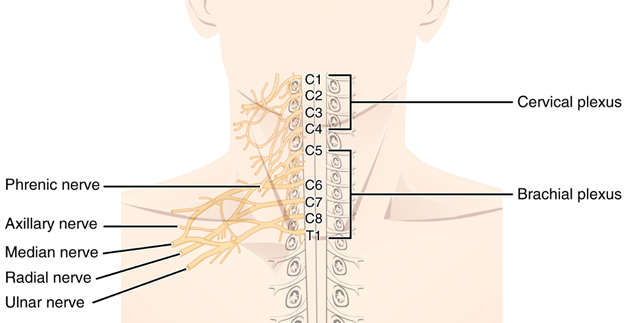
Figure 19.4—The cervical and brachial plexus in humans.
Notice that at first glance, the plexus is a mess of connections, splits, connections, splits, and a few more connections. With a complex structure like the brachial plexus, it helps to define regions rather than individual elements until you get to the major nerves themselves. The other major plexus across tetrapods is found near the pelvis, associated with the hindlimbs. One unique element to note is that the spinal nerves are named for their location along the vertebral column. In the mammals, the vertebral column is divided into regions (Chapter 9), but outside of the mammals this is not the case. When investigating the brachial plexus of an alligator, for example, you would note it as originating from spinal nerves VII–XI.
19.5 Evolution of the Cranial Nerves
Before we begin to explore the evolution of the cranial nerves, we should take a moment to remind ourselves of what the cranial nerves are and why they are important both in the context of evolution as well as being diagnostically important in both human and veterinary medicine. The cranial nerves function as spinal nerves do—individual neurons that carry signals to and from the brain. As was the case in the spinal nerves, they may only carry a signal in one direction, but most of the cranial nerves are intermixed, carrying both sensory and motor neurons. There are a few cranial nerves that are entirely sensory or entirely motor, and we will point these out when we encounter them. What makes them diagnostically important is how they interact with the CNS. The cranial nerves are special in that they connect directly to the brain. In a diagnostic setting, this means that disruption in their function can pinpoint damage to the brain itself without having to trace a path that includes other nerves and the spinal cord. They are also important because they are the nerves for the major external senses: vision, smell (olfaction), taste, hearing, and balance. Their evolutionary history also tells the tale of changing environments, historical functions, and how evolution tweaks existing structures through time.
As a matter of housekeeping before we dive in, we will agree to a common nomenclature when we refer to the cranial nerves. All the cranial nerves are given a name and number. The code “CN” is used in front of a Roman numeral with the name. For example, the nerve that runs from the eye to the brain is known as the optic nerve, and we will use CN II or refer to it by name. The cranial nerves number 0–XII, numbered in order from anterior to posterior (Figure 19.5).
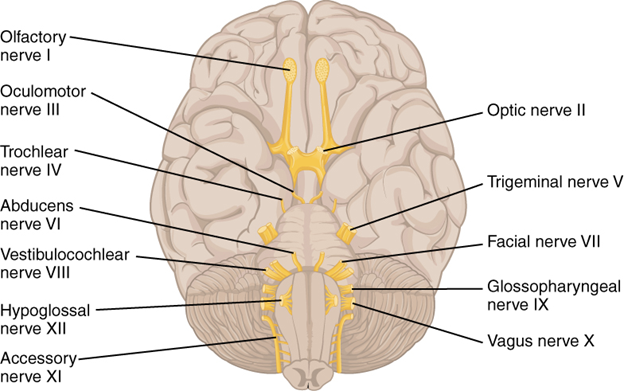
Figure 19.5—The human cranial nerves.
We will approach this section a little differently from the other chapters in this book. Instead of our straightforward march through the major vertebrate groups, we will instead introduce each nerve and its primary function. There will be some discussion as to whether it is a primarily sensory nerve or motor nerve or contains elements of both. From there, we will work through our vertebrate groups in order to better understand how the changes in some of the cranial nerves reflect much larger changes to many of the other systems involved in the anatomy of the head and neck.
Cranial Nerve 0: The Terminal Nerve
The first cranial nerve is special in a few ways. First, it was only discovered and officially named after most of the other cranial nerves (hence it being referred to as CN 0). Second, it is only grossly visible in a few groups of fishes. However, its presence in other vertebrates (including humans) has been known for some time. The function of the terminal nerve is still unclear. It originates in the brain near the olfactory centers and has been observed to be active in correlation with sex hormone activity. With this limited evidence, it is currently presumed to play a role in olfaction, particularly of pheromones as they relate to reproduction. It originates on the cerebrum before heading anteriorly.
Cranial Nerve I: The Olfactory Nerve
The olfactory nerve (CN I) is a great nerve to get started on the deep dive of the cranial nerves (ignoring the terminal nerve, like so many anatomists before us). As its name implies, the olfactory nerve delivers sensory information from the olfactory epithelium in the nasal passages to the brain. Unlike many of the other cranial nerves, the olfactory nerve carries only information to the CNS. In this way, we may say that the olfactory nerve is sensory only. The olfactory nerve carries information about odor, what we know as the sense of smell, to the brain. Depending on the particular vertebrate group, that may be a proper olfactory bulb or tract before diving deeper into the brain. The olfactory nerve originates on the cerebrum, associated with structures known as olfactory bulbs.
Cranial Nerve II: The Optic Nerve
The optic nerve (CN II) is also a purely sensory nerve, carrying visual information from the retina to the optic centers of the brain from its origin in the cerebrum. These paired nerves cross/meet each other at the optic chiasma before continuing on to the optic lobes.
Cranial Nerve III: Oculomotor Nerve
The oculomotor nerve (CN III) is the most anterior nerve to originate from the midbrain. Unlike the previous cranial nerves, the oculomotor (as the name suggests) is motor only. In many vertebrates, there are several skeletal muscles that control the movement of the eye itself, known as extraocular muscles. Most, but not all, of these muscles are controlled by the oculomotor nerve.
Cranial Nerve IV: Trochlear Nerve
The trochlear nerve (CN IV) is also a motor-only cranial nerve that also controls eye movement, originating in the midbrain. However, unlike the oculomotor nerve, the trochlear nerve controls only one muscle, a muscle that in humans we refer to as the superior oblique muscle. The superior oblique muscle has an odd course, anatomically speaking. It starts similarly to the other extraocular muscles, but its tendon passes through a cartilaginous pulley before inserting on the lateral aspect of the eye. This small pulley is known as a trochlea (Latin for “pulley”). This is where the trochlear nerve gets its name.
Cranial Nerve V: Trigeminal Nerve
The trigeminal nerve (CN V) is one of the largest cranial nerves, with a sizable and noticeable origin on the brain stem at the level of the pons. The trigeminal nerve (tri-, Latin for “three,” and -geminus, Latin for “twin”) is known for its three major branches that all course through the head: the ophthalmic branch (commonly denoted as V1), maxillary branch (V2), and mandibular branch (V3). With its large branches and huge area of innervation, it should be no surprise that the trigeminal nerve is our first mixed nerve. This means that it has both sensory and motor components. The trigeminal nerve is very important for several sensory components in the head (outside of olfaction and vision) and is the main innervator of the jaw-closing muscles. We will explore the trigeminal nerve in detail through our exploration of the evolutionary history of vertebrates, as it moves and changes frequently through time. We will also spend a lot of time with this nerve as a diagnostic tool for human health.
Cranial Nerve VI: Abducens Nerve
The abducens nerve (CN VI) is a nerve quite similar to CN IV except that it originates on the brain stem. It is a motor-only nerve, and it controls the last of the extraocular muscles we will consider here. Known as the lateral rectus muscle, it helps pull the eye laterally. This nerve follows a similar course and has the same function in all the vertebrates with the exception of the hagfish and lamprey, where it is seemingly absent.
Cranial Nerve VII: Facial Nerve
The facial nerve (CN VII) is another one of the cranial nerves that we will revisit frequently as we explore the different vertebrate groups. The facial nerve is a mixed nerve, with both sensory and motor components. Do not be fooled by the name, as it does not originate near the face (it is also in the brain stem) and its actions are also more complicated. The facial nerve only innervates muscles and sensory receptors in the face of mammals. In most of our other vertebrates, we will see that the facial nerve performs many other tasks in other regions of the head and neck. In many ways, the facial nerve is a great example of how evolution tweaks and tinkers with existing structures. Following the track of the facial nerve, we can see quite clearly how changes in structure and function in some elements of anatomy have corresponding changes downstream. It is also a great example to see how the transition from water to land impacted not only the “major” musculoskeletal structures associated with movement and feeding but also the supporting structures. The facial nerve is also diagnostically important in humans, with many diseases and tests relying on the facial nerve.
Cranial Nerve VIII: Vestibulocochlear Nerve
The vestibulocochlear nerve (CN VIII), sometimes also referred to as the statoacoustic or auditory vestibular nerve, is a mixed nerve playing a role in hearing and balance, originating on the brain stem. These two senses are closely associated with each other in many vertebrates (see Chapter 20—Nervous System: Sensory Organs). The vestibulocochlear nerve is quickly separated into the vestibular (balance) and cochlear (hearing) divisions. Hearing and balance vary greatly in vertebrates but do exist across almost every group. While technically a mixed nerve, the motor component of CN VIII is rather minimal. In humans, the motor components are involved in a reflex involving the vestibular system and the eyes, with the vestibulocochlear nerve innervating the extraocular muscles.
Cranial Nerve IX: Glossopharyngeal Nerve
The glossopharyngeal nerve (CN IX) is much like the vestibulocochlear nerve in that its primary functions can be determined by digging into its name. Glosso- often refers to tongues, coming from a Greek origin (“tongue, word, speech”), while the second half of the word is derived from the pharynx. There are many pharyngeal muscles, and this nerve plays a role in innervating many of those muscles. The glossopharyngeal nerve is another mixed nerve originating on the brain stem, with motor neurons for several muscles in the head and neck region (including the tongue in those vertebrates that have one). Sensory information comes from many different sources, including taste from the tongue, chemical sensing from several blood vessels, and information from the pharynx.
Cranial Nerve X: Vagus Nerve
The vagus nerve (CN X) is one of the longest nerves in the body. Elements of the vagus nerve innervate structures from the head all the way down to the abdominal organs. The vagus is also a mixed nerve; however, it is important to note some of the key differences that make the vagus unique (especially in humans). To this point, we have mostly been discussing the motor aspect of cranial nerves as controlling different skeletal muscles. However, as you will know from Chapter 11 (muscles), not all muscle in a body is skeletal. Cardiac muscle and smooth muscle still contract and still need at least some element of nervous system control (cardiac muscle will contract on its own but is regulated by the nervous system). Many of the tubular organs, with smooth muscle, are regulated by the vagus nerve. Heart rate is regulated through the vagus nerve (and the endocrine system), but there is still some controversy over the role of the vagus nerve in the force of heart muscle contraction. The vagus is also unique in that for several parts of its journey down the body, it contains both sympathetic and parasympathetic nerve fibers along its path. Along the neck and entering the chest, we refer to this conjunction as the vagosympathetic trunk, before the elements of the sympathetic trunk split off.
Cranial Nerve XI: Accessory Nerve
The accessory nerve (CN XI) is an interesting case in the world of the cranial nerves. The accessory nerve is a motor-only nerve, providing control of several muscles along the back of the neck and pectoral girdle. What makes the accessory nerve interesting is that it does not entirely originate from the brain. The accessory nerve has several rootlets that originate on the spinal cord. In mammals, these rootlets join together and travel cranially. They make a loop and reexit the skull, and this is why it was named as a cranial nerve. I hate the accessory nerve.
Cranial Nerve XII: Hypoglossal Nerve
The last official cranial nerve, the hypoglossal nerve (CN XII), originates on the medulla oblongata and immediately exits the skull. It is a motor-only nerve, and it also controls several of the muscles associated with the tongue.
Now with introductions out of the way, let us see how these nerves change through evolutionary time in response to changing structures, habitats, and functions.
Cyclostomes
While representing the earliest diverging lineage of vertebrates, it is beneficial to remind ourselves that both the hagfish and lamprey are highly specialized evolutionary lineages in their own rights. So while they are, in other body systems, used to represent the ancestral condition, they can show some specializations that may confuse the investigator. In the lamprey, all 12 pairs of cranial nerves can be located and distinguished, and the presence of a terminal nerve is still contested. Hagfishes also seem to have all 12 nerves, but some recent work finds that CN X–XII are hypermodified in the hagfish and that CN XI and XII may actually be independent evolutions of those nerves rather than the result of shared ancestry.
Fishes
In most comparative anatomy resources, the cranial nerves of sharks serve as the de facto representative of the vertebrate ancestral condition (Figure 19.6). Fortunately for us, there are not a lot of differences between the different groups of fishes. There is some variation naturally, but much of it is beyond the scope of this book. With that in mind, we will focus on the sharks, noting key differences when they appear. Eleven pairs of cranial nerves can be found in sharks, missing CN XI, the accessory nerve. All the cranial nerves remain separate from each other until they reach their destination, unlike the hagfish and lamprey. Because they remain separate from each other, it is also easier to determine what their function is. For the first few cranial nerves, we see that the cartilaginous fishes set the trend that the other vertebrates will follow.
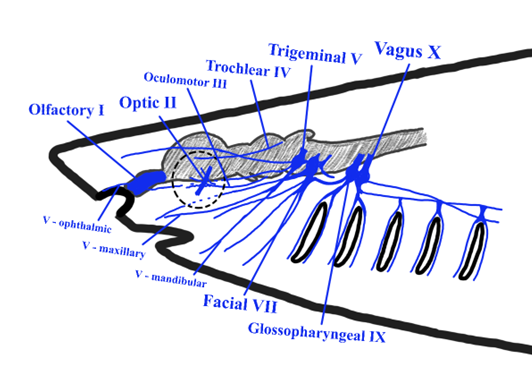
Figure 19.6—The cranial nerves of a generalized shark. Not visible are terminal (CN 0), abducens (CN VI), and vestibulocochlear (CN VIII) nerves.
The terminal and olfactory nerves are closely associated with each other. The olfactory nerves are quite short, running to the olfactory bulbs. The terminal nerve is a much longer nerve, running from the hypothalamus section of the brain to the nares. As we stated earlier, the terminal nerve plays some role in olfaction, currently hypothesized to be involved in the detection of pheromones, but evidence is scant. The optic (CN II), oculomotor (CN III), trochlear (CN IV), and abducens (CN VI) nerves serve the same function as in other vertebrates, which we discussed earlier. The first nerve that differs from the general pattern is the trigeminal nerve (CN V). The trigeminal nerve still has four major branches in sharks, with the previously mentioned ophthalmic branch being separated into the ophthalmicus profundus and ophthalmicus superficialis. The mandibular branch is subdivided into superior and inferior branches. The main sensory function of the trigeminal in sharks is to receive sensory information from the skin around the snout. However, there is an important distinction to make here. The ampullae of Lorenzini, electroreceptors in the snout of most cartilaginous fishes, are not innervated by the trigeminal nerve. The nerves that carry the signals from the electroreceptors are derived from similar nerves for the lateral line system. These nerves are technically cranial nerves, as they go directly to the brain, but they are not given official names or numbers. The trigeminal’s motor nerves innervate the jaw-closing muscles: adductor mandibularis being the major jaw closer in sharks.
The facial nerve (CN VII) reveals its mammalian naming bias in the cartilaginous fishes. The facial nerve…has nothing to do with the face. It was originally thought to receive sensory information from the most anterior portions of the lateral line system, though it has been determined in several shark species that those are independent nerves that run near the facial nerve. However, the facial nerve does still play some role in the sensory system of sharks, receiving information from taste receptors scattered throughout the mouth. The motor components of the facial nerve are split between areas associated with the head and the anterior body. Within the head, the facial nerve controls several of the pharyngeal muscles, those associated with the second pharyngeal arch. It also contributes to the control of muscles and mucus membranes associated with the first gill. If you check out the muscles chapter (Chapter 11), you will see these muscles make some pretty big migrations as vertebrates moved from an aquatic environment to a terrestrial one, and the facial nerve travels along with those muscles. This will also have some implications for the role of the facial nerve in human health (Box 19.1). The statoacoustic nerve (CN VIII), as it is often referred to in the fishes, does innervate the hearing and balance sensory receptors. In addition, it also has efferent innervation of the lateral line system. This does not mean it also receives information from the lateral line system. In fact, the lateral line system is innervated by two unofficial cranial nerves, the anterior lateral line nerve and the posterior lateral line nerve. The glossopharyngeal nerve (CN IX) represents another mammalian bias in the naming of the cranial nerves. In our fishes, CN IX plays a larger role in the first gill arch (third branchial arch). Fishes lack a true tongue, and the glossopharyngeal nerve, given its position, is more directly associated with the first few gills than the true pharynx. The last officially recognized cranial nerve in fishes is CN X, the vagus nerve. In our general introduction to the cranial nerves, we indicated that the vagus nerve serves a lot of important roles in the body, especially beyond the head. In the shark, the vagus is a very large nerve. Its impacts are widespread, from the last few gills to almost the cloaca itself. The vagus impacts the esophagus, heart, stomach, intestine, and parts of the body wall itself.
Tetrapods: A Brief Note
With the movement of the tetrapods from the water to the land, there were a few changes in the spinal nerves as limbs evolved and became the driving force for locomotion. However, these changes could be generalized across the larger groups of tetrapods. As we make our way into the cranial nerves of these same groups, we will find that this is not going to be the case. To further a theme already described in other chapters (but take a look at the skull and muscle chapters [Chapter 8 and Chapter 1] in particular), the transition from water to land caused a great deal of changes to the sensory systems and feeding apparatus of tetrapods. It should not be surprising then that the cranial nerves associated with those systems are the ones that we are going to observe the most changes in. In our remaining tour of the vertebrates, we will focus on those changes. CN I–IV and VI will largely go unmentioned, as they continue to function in the same ways as we described in the generalized section. The unnamed lateral line nerves will reduce and disappear. CN X continues to dominate control of the body wall and the visceral organs, but changes to its innervation cranially will require some attention. The nerves not already mentioned must be considered in detail, and we will find some surprising trends among all this variation.
Nonmammalian Tetrapods
There are two seemingly innocuous changes (and one major one) that happen in the head as you move onto land that impact the skeleton, muscles, and therefore the cranial nerves. These are the evolution of the tongue and neck. The big change, the loss of the gills, is obviously going to impact a lot of these structures as well. However, many of the cranial nerves undergo little or no change as we transition to a fully terrestrial lifestyle. CN II, III, IV, VI, and VIII are quite similar in the tetrapods to their condition in the fishes. As will be discussed briefly in the sensory systems chapter (Chapter 20), there is an additional or accessory olfactory organ that appears and is particularly well developed in the reptiles (and mammals) known as the vomeronasal organ, or VNO for short. The olfactory nerve (CN I) serves this accessory organ, and in some groups the individual nerves that innervate the vomeronasal organ form a distinct branch. The trigeminal nerve (CN V) is more variable in the reptiles than it would seem to be in the other vertebrates. This is in large part due to the evolution of a few “new” senses and their distribution in the head. Alligators and crocodiles have unique touch receptors embedded in and around the scales of their jaws and snouts called integumentary sense organs (ISOs). These ISOs are innervated by the major branches of the trigeminal nerve. The same can be said for the infrared “vision” of the vipers, boas, and pythons. In these snakes, the path of the trigeminal nerve takes a detour. CN V in these snakes will have several small nerves move toward the visual processing centers of the brain, which is why we refer to this infrared reception as a type of vision. The trigeminal nerve is associated with the jaw closers, and that remains true. But it will also be part of that new structure in the mouth, the tongue. The muscles of the tongue are not derived from the jaw closers, instead originating from farther back. However, some of the sensory components of the tongue (not taste) do send their signals to the brain along the trigeminal nerve. Conversely, the venom glands of snakes have unique muscles that control venom output during a bite. These muscles are controlled by the trigeminal nerve. Beyond the trigeminal nerve, there is more considerable variation. The facial nerve (CN VII) is highly variable, even within groups. Recall that the facial nerve in fishes innervated elements of the hyoid/pharyngeal muscles as well as the first gill arch. For our fully aquatic salamanders (like the mudpuppy), this is still the case. However, in our terrestrial amphibians and our reptiles, the changes may be pretty dramatic. It may be helpful to think of the facial nerve in the context of what is happening to the muscles in this area. As the muscles develop around the second and third visceral arch, many of them migrate cranially. The facial nerve travels with them. Part of the tongue is innervated by CN VII, and this is where the sense of taste comes in. The other brand-new element in the head is the appearance of salivary glands. The salivary glands help in the chemical digestion and transport of food along the digestive tract. They are also linked to the nervous system, which can control output and when salivation occurs. These new glands are innervated by the facial nerve.
The next really big changes occur when we reach CN IX, the glossopharyngeal nerve. In our fishes the glossopharyngeal nerve, despite its name, was confined to the gills, where it played a large role in controlling the muscles associated with the gills. Once again, this does not change in our aquatic salamanders. With the transition to land, the gills disappear. What happens to all those muscles; do they disappear as well? We know that this is not the case. Instead, the muscles migrate cranially, becoming part of the musculature of the pharynx and the majority of the tongue. The glossopharyngeal nerve travels with the muscles as they migrate and becomes one of the major nerves that control the muscles of the tongue. In our fishes, we moved on to the vagus nerve (CN X), and that was the end. The vagus nerve in fishes we said interacted with the more caudal gills and their muscles, plus all the internal organs in the body cavity. The internal organs in our tetrapods still have a lot of vagal innervation, but it is the fate of those nerves that control the gill muscles where changes happen. There is an important shift when we get to the tetrapods (particularly the reptiles for now). With the reduction in dermal bones (Chapter 11) and the evolution of a distinct neck, the function of CN X also changed.
The neck requires muscles to provide stability and allow for movement. While homologous to the gill muscles, the innervation pattern evolves in correlation with the muscles. We get the reemergence of the accessory nerve (CN XI). The accessory nerve has an interesting developmental and evolutionary tale to tell. In fishes, the accessory nerves originate beginning at the junction of the brain stem and spinal cord and a few spinal nerve rootlets. It travels a short distance and anteriorly, where it merges with the vagus nerve. As a result, many researchers do not consider the fishes to have a true accessory nerve. With the evolution of a neck region, the origin of the accessory nerve moves more posteriorly, splitting from the vagus nerve and becoming its own distinct cranial nerve. Developmentally, the accessory nerve isn’t a cranial nerve at all. It originates on the spinal cord, in roughly the sections corresponding to C1–C6. However, after it emerges, it heads cranially, enters the skull, and then promptly exits the skull again—technically a “cranial” nerve, as it leaves the cranium, but it does not originate from the brain. We will get into its specific pathway in humans, as there are some clinical implications of this movement. However, we cannot forget the vagus nerve and its role in the gill muscles. Given the original proximity of the gills to the pharynx, some of the branches of the vagus are associated with the pharyngeal muscles. This is still true, and branches also innervate the muscles of the larynx in our tetrapods. One branch of the vagus nerve, known as the recurrent laryngeal nerve, leaves the vagus in the thoracic (chest) cavity and makes its way all the way back into the neck before reaching some of those laryngeal muscles. Last, we arrive at our discussion of the hypoglossal nerve. The hypoglossal nerve (CN XII) emerges as a new nerve not described or found in the fishes and only in tetrapods. Developmental evidence suggests that it originated as a branch of the vagus nerve, as is hypothesized for the accessory nerve. As its name implies, the hypoglossal nerves course under the tongue and also innervate several of the muscles in the tongue. These muscles are critical for moving food within the mouth and swallowing.
Humans and Other Mammals
With a few notable exceptions, we can use human cranial nerves to describe all the functions of the cranial nerves in mammals. While exploring the patterns and functions of the cranial nerves, we will also discuss their clinical relevance in humans (Figure 19.6).
The presence of a terminal nerve in mammals is still being hotly debated. While there is no doubt that some mammals communicate with pheromones, it remains unclear if the terminal remains as an independent nerve or has become a branch of the olfactory nerve (CN I). The olfactory nerve is very short in mammals, extending only a few millimeters through openings in the skull known as the cribiform foramina. In humans, damage to the olfactory nerve results in the loss of smell. Cranial nerves II–IV and VI (optic, oculomotor, trochlear, abducens) function in mammals as they do in other vertebrates. The optic nerve is responsible for vision. To test for damage in the optic nerve, asking someone if they are able to see is not enough. Recall from the brain chapter (Chapter 18) that the visual processing centers in humans are in the rear of the brain. How do you distinguish damage to the visual processing centers from damage to the physical structures of the eye? This is where the pupillary reflex comes in. Shining a bright light into the eyes results in contraction of the muscles of the iris, shrinking the pupil. This reflex does not require input from the visual processing center and can be used to test for damage in the optic nerve. The other three we know control the muscles that move the eyeball within the orbit. You can test these nerves yourself. Hold your finger in front of your face and keep your head perfectly still. Now move your finger to your left, right, up, and down. Was your eye able to track your finger in each direction? What if you move it diagonally? Look at the tip of your nose? If so, congratulations, those four cranial nerves (optic, oculomotor, trochlear, and abducens) are all intact. Damage to any of those nerves results in difficulty tracking or even maintaining a straightforward gaze. The eye may “roll” in a direction even at rest with damage to the trochlear or abducens nerves.
The trigeminal nerve, as we already know, is a large and important nerve for all vertebrates. It still controls the jaw-closing muscles, as well as getting considerable sensory feedback from the many elements of the head. An excellent example of this can be found in the platypus. The platypus has electroreceptors that can detect small electric fields in water. The signals from these receptors are carried on the trigeminal nerve. In humans, damage to the trigeminal nerve can manifest itself in many different ways. Difficulty chewing, speaking, and swallowing can all be related to trigeminal nerve damage. Simultaneously, because of the amount of sensory information being carried by the trigeminal nerve, numbness in particular parts of the face and mouth points to the trigeminal nerve. If you’ve had a cavity filled or your wisdom teeth removed, you know this all too well. The superior and inferior alveolar nerves innervate the teeth of the upper and lower jaws, respectively. Both nerves are branches of the trigeminal nerve, coming from the maxillary and mandibular branches.
The facial nerve (CN VII) in humans and other mammals has also changed since we last visited it in the nonmammalian tetrapods. When we discussed amphibians and reptiles, the facial nerve played a large role in the tongue and hyoid apparatus. However, as mammals evolved, there were different changes to those muscles (recall that reptiles and mammals share a common ancestor from a long, long time ago). Much of the musculature, developmentally, that originates in the pharyngeal region moves to cover the face in mammals. We call these muscles the muscles of facial expression. Smile, furrow your brow, pretend to be shocked. A lot of that movement was directed by the facial nerve. The tortuous route of the facial nerve, beginning behind the ear in humans and working its way to the face, has some diagnostically important implications, sometimes resulting in paralysis of half the face (Box 19.1). Similarly to amphibians and reptiles, the facial nerve also stimulates salivary gland activity and is the home to our sense of taste. Excessive salivation or the loss of taste may independently indicate damage to the facial nerve.
Box 19.1—The Facial Nerve, Bell’s Palsy, and Ear Infections
The facial nerve serves as a good reminder that the cranial nerves are paired, with one coming off the left and one coming off the right side of the brain. A loss of one of the nerves may only impair that side of the body. This is the case for Bell’s palsy, the name of the condition where one of the facial nerves is damaged, resulting in paralysis to that half of the face. The causes of Bell’s palsy are myriad, but there is one that should draw our attention in the context of the evolution of vertebrates. Draw the path of the facial nerve from its origin to all those facial muscles. You notice that it passes right around the paths of the middle and inner ear. Why does this matter? In severe ear infections, the pressure buildup in the ear will compress the facial nerve, resulting in the paralysis of the facial muscles! Going from a gill muscle controller to tongue and hyoid nerve to the muscle of facial expression has twisted and turned the facial nerve in all sorts of directions, and not always for the better.
The vestibulocochlear nerve (CN VIII) carries the information for our senses of hearing and balance. Hearing tests that include pitch, volume, and direction can all be used to determine function of the vestibulocochlear nerve. Additionally, you may test balance by standing on one foot with the other pointed forward. It is important, when testing hearing, to test both sides of the head to determine whether damage is localized to one of the nerves or to the brain itself.
The glossopharyngeal nerve (CN IX) is very important in the movement of the tongue and pharynx. It is also the key player in the gag reflex. This combination of swallowing and gagging means that while it is clearly of functional importance, it is also an easy nerve to test for function. Loss of the gag reflex may reflect damage to CN IX (if you’re planning on testing this reflex, we recommend keeping a bucket nearby just in case). Beyond its motor components, the glossopharyngeal nerve serves critical sensory functions. The glossopharyngeal nerve is closely associated with the carotid arteries, and it is here that blood pressure and blood oxygen can be monitored.
The vagus nerve (CN X) is of critical importance in human health. As in other vertebrates, the vagus innervates some elements of the pharynx, larynx, and internal organs. The vagus directly interacts with the heart, lungs, and digestive tract. The vagus can impact heart rate, breathing rate, and the movement of food through the digestive system. Additionally, it receives information from blood pressure sensors and position sensors in the larynx. The recurrent laryngeal nerve is quite long in people and even longer in some mammals (Box 19.2). Diseases and disorders involving the vagus nerve are prevalent throughout the medical and veterinary community (see 19.7 Further Reading). As the vagus plays a role in swallowing, it can be tested similarly to the glossopharyngeal nerve.
Box 19.2—Longest Nerve in the World, the Recurrent Laryngeal Nerve
The recurrent laryngeal nerve is a branch of the vagus nerve that innervates several of the muscles that are important for swallowing and producing vocal sounds. Why are we dedicating a whole box to just one nerve? The recurrent laryngeal nerve is a great example of how development and evolution tinker with existing structures, with the result not always making the most sense. In sharks and other fishes, there is no recurrent laryngeal nerve; instead, several of the branches of the vagus nerve innervate the gill muscles (Figure 19.7A). The evolution of the tetrapods changes both the musculature in this region of the body (Chapter 11) as well as the fate of the aortic arches (Chapter 15). Some of these muscles become those of the larynx. The heart and aortic arches move caudally in time, and those muscles stay close to the head. The recurrent laryngeal gets caught by the ligamentum arteriosum (the remnant of a shunt between the aorta and pulmonary trunk), wrapped around the remaining aortic arches but still needing to reach the neck, as seen in the mouse (Figure 19.7B). The most striking example of this can be seen in the giraffe (Figure 19.7C). In giraffes, the left recurrent laryngeal nerve takes a detour that is over 4.5 meters (15 feet) in length!
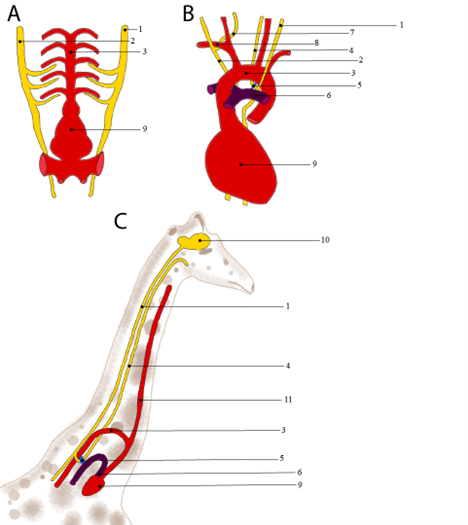
Figure 19.7—The evolution of the recurrent laryngeal nerve from the ancestral condition in the shark (A) to the mouse (B) and its extreme in the giraffe (C). Labeled structures are (1) left vagus nerve; (2) right vagus nerve; (3) aorta; (4) left recurrent laryngeal nerve; (5) ligamentum arteriosum; (6) pulmonary trunk; (7) right recurrent laryngeal nerve; (8) right subclavian artery; (9) heart; (10) brain; and (11) common carotid artery.
The accessory nerve (CN XI) is entirely motor in humans, innervating the trapezius and sternocleidomastoid muscles. The movements of these large muscles are obvious to an observer, and any difficulty in their movement should point to damage in the nerves. Much like in reptiles, the accessory nerves actually begin in the spinal cord. Many spinal rootlets merge together before entering the skull via the foramen magnum. After entering, the accessory nerve makes an about-face, exiting the skull via the jugular foramen on its way down to the muscles.
Finally, the hypoglossal nerve, CN XII: As was the case in the reptiles, the hypoglossal nerve innervates many of the muscles of the tongue. The tongue muscles controlled by the hypoglossal nerve are of critical importance not only to swallowing but to speech as well. Speech is an incredibly complex process, involving several areas of the brain. Therefore, relying on speech as a diagnostic tool is not particularly helpful in the context of the hypoglossal nerve. Instead, a small motor test can reliably isolate the function of the hypoglossal nerve. If you can stick your tongue out, both protracting and retracting the tongue, then the function of the hypoglossal nerve is intact.
19.6 Summary
The peripheral nervous system is the link between the central nervous system and the rest of the body. It carries sensory information to the brain and outgoing effector information to muscles, glands, and organs that respond to changes. Some of these nerves, the cranial nerves, bypass the spinal cord, communicating directly with the brain and making them invaluable structures for understanding vertebrate evolution as well as clinically relevant human anatomy.
Application Questions
- How do reflexes act as a miniature version of the nervous system itself?
- Why does damage to one optic nerve not result in blindness in that eye?
- How can some nerves be considered mixed?
19.7 Further Reading
- The Developing Cranial Nerves in Zebrafish
- Vasovagal Syncope and Laryngeal Hemiplegia in Horses
- The Development of the Cranial Nerves
19.8 References
- Davis, Matthew C., Christoph J. Griessenauer, Anand N. Bosmia, R. Shane Tubbs, and Mohammadali M. Shoja. “The naming of the cranial nerves: A historical review.” Clinical Anatomy 27, no. 1 (2014): 14–19.
- George, Ian D., and Casey M. Holliday. “Trigeminal nerve morphology in Alligator mississippiensis and its significance for crocodyliform facial sensation and evolution.” The Anatomical Record 296, no. 4 (2013): 670–680.
- Ito, Hironobu, Masami Yoshimoto, and Hiroaki Somiya. “External brain form and cranial nerves of the megamouth shark, Megachasma pelagios.” Copeia (1999): 210–213.
- Lessner, Emily J., and Casey M. Holliday. “A 3D ontogenetic atlas of Alligator mississippiensis cranial nerves and their significance for comparative neurology of reptiles.” The Anatomical Record 305, no. 10 (2022): 2854–2882.
- Mallatt, Jon. “Shark pharyngeal muscles and early vertebrate evolution.” Acta Zoologica 78, no. 4 (1997): 279–294.
- Northcutt, R. Glenn. “Elasmobranch central nervous system organization and its possible evolutionary significance.” American Zoologist 17, no. 2 (1977): 411–429.
- Oisi, Yasuhiro, Satoko Fujimoto, Kinya G. Ota, and Shigeru Kuratani. “On the peculiar morphology and development of the hypoglossal, glossopharyngeal and vagus nerves and hypobranchial muscles in the hagfish.” Zoological Letters 1 (2015): 1–15.
- Pombal, Manuel A., and Manuel Megías. “Development and functional organization of the cranial nerves in lampreys.” The Anatomical Record 302, no. 3 (2019): 512–539.
- Rea, Paul. Clinical Anatomy of the Cranial Nerves. Academic Press, 2014.
- Ruqa, Wael Abu, Fiorenza Pennacchia, Eqrem Rusi, Federica Zoccali, Giuseppe Bruno, Giuseppina Talarico, Christian Barbato, and Antonio Minni. “Smelling TNT: Trends of the terminal nerve.” International Journal of Molecular Sciences 25, no. 7 (2024): 3920.
- Vilensky, Joel A. “The neglected cranial nerve: Nervus terminalis (cranial nerve N).” Clinical Anatomy 27, no. 1 (2014): 46–53.
