
7 Bone and Cartilage
Bill Ryerson
Focus Questions—to Guide Your Reading of This Chapter
- What do the prefixes and suffixes of cell names tell us about the connective tissues?
- Cells are important in every system, but it seems that everything outside of the cells in bones and cartilage is stressed in this chapter. Why might that be?
- How do the different types of bone and cartilage and their structural properties impact where we find them in the skeleton?
7.1 Introduction
For the next several chapters, we will take a deep dive into the skeleton. Like the previous chapter on the integument, we will go through a generalized introduction to the different elements of the skeleton, where they originate developmentally, and how their variation across vertebrates can tell us about the evolutionary history and functional importance of those structures. The skeleton is such a broad system, with so many different moving pieces (pun very much intended), that it would make sense to talk about the components of the skeleton first, how they function, and where they come from before diving into the particular regions.
All vertebrates possess an endoskeleton—that is, a skeleton that is internal to most of the body (endo- from the Greek endon, meaning “within”). This is somewhat of a vertebrate-defining characteristic, as few other animals possess a rigid internal skeleton. Arthropods, the largest group of animals on the planet (insects, spiders, lobsters, etc.), possess an exoskeleton. There may be skeletal elements that are on the external surface of a vertebrate (see the osteoderms in Chapter 6), but the major pieces of the skeleton are internal to the body. The skeleton may be highly reduced to only a few elements, or it may have hundreds of individual elements. It can be made of bone, as it is in humans, or it can be predominantly cartilage, as in our cartilaginous fishes. There are even some gray areas, where cartilage can look and act like bone, as we see in some of our larger sharks. We will discuss the details of cartilage and bone, how they develop, and their functional roles later in this chapter.
7.2 What Is the Skeleton?
As we just discussed, the skeleton is a primarily internal structure of tough tissues. To really think about the skeleton, we must consider what it does in vertebrates to reflect on how structural variability links with function. We must also consider some of the lesser-known functions of the skeleton, as they may help us understand some of the constraints or limits to the structural variability we see in the skeleton. Skeleton functions include the following:
- Provide support for the body
- Protection of internal organs (think about the brain or heart)
- Anchors for muscles to provide movement (muscles create movement, but the bones actually move)
- Mineral/nutrient storage
- Formation of new blood cells—white and red
- Vitamin synthesis
When we consider the skeleton, we will focus primarily on the first three functions, as they will help us understand the evolution of the skeleton as well as how it impacts the evolutionary history of the vertebrates. However, do not neglect the last three functions, as they do impact the structures of the skeleton and play a vital role in human health.
Despite the fact that the skeleton itself can be thought of as a single system, almost all anatomy textbooks and references (and anatomists themselves!) divide the skeleton itself into a few regions. Some of this is functional in nature, some developmental, and some evolutionary. We will see that the skeleton as well as individual bones and joints will be categorized according to similar patterns, and once we make those patterns obvious, the distinctions will hopefully fall into place.
The first skeletal dichotomy is the axial skeleton versus the appendicular skeleton (Figure 7.1).
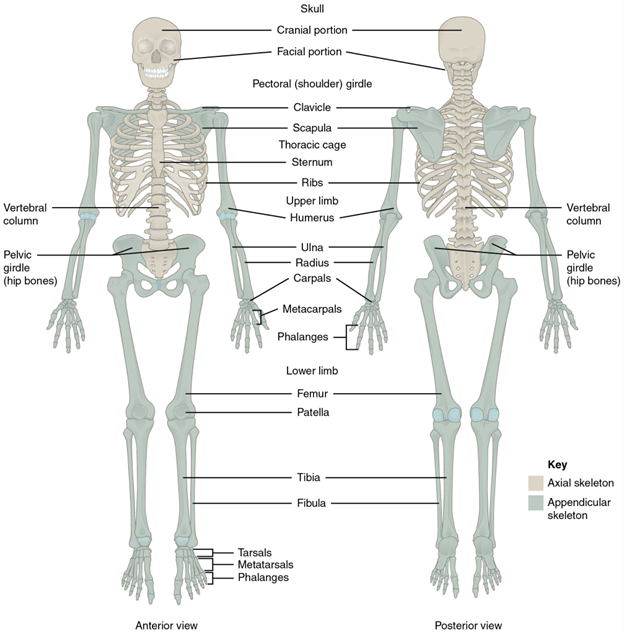
Figure 7.1—Axial and appendicular skeleton. The axial skeleton supports the head, neck, back, and chest and thus forms the vertical axis of the body. It consists of the skull, vertebral column (including the sacrum and coccyx), and thoracic cage, formed by the ribs and sternum. The appendicular skeleton is made up of all bones of the upper and lower limbs.
Take the human body and make a vertical cut right down the midline (gross), from head to feet. All the bones you would cut through, with two notable exceptions, are part of the axial skeleton. The exceptions are the pelvis (part of the appendicular skeleton) and the ribs, which while you did not cut through them, are part of the axial skeleton. Evolutionarily, these are the “original skeleton.” The bones associated with the appendicular skeleton are all the other bones of the body. Their name hopefully gives them away, as they are the bones that either are part of the appendages or anchor the appendages to the axial skeleton.
The second dichotomy is the cranial versus the postcranial skeleton. Hopefully this one is even more straightforward than the previous. The “cranial” skeleton is the skull and its associated structures, like the hyoid. This is both a developmental and evolutionary distinction. As we will see in Chapter 8, the skull has a long and storied history, and so many will focus on the skull, leaving the rest of the skeleton on its own as postcranial (see Chapters 9 and 10).
The third dichotomy is dependent on the developmental formation of the bones. The two types are endochondral and intramembranous bones. These refer to the origin of those bones during development. The process of forming bones in the skeleton is known as ossification. Future hint: Wherever you see “oss-,” it is referring to bone or something analogous to bone. It is not always bone, as many large sharks and other cartilaginous fish will ossify parts of their jaws, skulls, and vertebrae, which are different from true bone. Later in this chapter, we will distinguish the differences between cartilage and bone. When an embryo is developing, not all bones form in the same way. Some start off as cartilage molds, which are gradually replaced by bone. These are the endochondral bones (endo = within; chondra = cartilage). Other bones will form within developing sheets of connective tissue. These are the intramembranous bones. Before we talk about how these bones will arise, let’s take a big step back and introduce the players that will guide these changes.
Bone and cartilage are alike in that they are both connective tissues. This means that they share several characteristics and that the differences among these characteristics will help us define the different tissues. Like any tissue type, there are cells. We can define the different bone and cartilage cells by their functions in the body and assign them names based on these functions. We will also define them as “immature” and “mature” depending on where they are in the life cycle of a cell. There are many types of connective tissues, but for this section we are going to be focusing on just bone and cartilage (and maybe one/two more). We can look at the names given to these cells and break them apart into their prefix and suffix. The prefix will tell us what tissue type that cell belongs to: osteo- for bone and chondro- for cartilage. The suffix will tell us the function and maturity of the cell: -blast for immature cells and -cyte for mature cells. So an osteoblast is an immature bone cell. A chondrocyte is a mature cartilage cell. Immature cells, for both bone and cartilage, are usually what we refer to as “builders.” Builders will create the material that ultimately composes bone and/or cartilage. Mature cells are often referred to as “monitors.” These mature cells will remain embedded within the tissue, checking for damage or changes in the chemical environment. They can send their own signals to indicate that the tissue needs to be repaired. We have osteoblasts, osteocytes, chondroblasts, and chondrocytes. There is one more cell type to be introduced for now. Bone has a third type of cell, the osteoclast. Clast has a Greek origin, meaning “to break.” Osteoclasts are cells that will break down bone for reasons that we will discuss later. Bone is a very dynamic tissue, always being broken down and replaced depending on activity, age, nutrition, and general health.
While the cells are important, they do not help us define what bone and cartilage are and how they function. All connective tissues have an additional component known as the extracellular matrix (ECM). The ECM is just as important as (if not more so than) the cells. The ECM is built from two major elements: fibers and a ground substance. The fibers are typically one of three types: collagen, elastin, and/or reticular fibers. Collagen is the most common protein in the body, found in muscle, bone, cartilage, tendons, ligaments, and more. Bone will have collagen, but some cartilages may have both collagen and elastin. The type of fiber and the amount will determine the flexibility and strength of a material. Collagen is the least flexible but the strongest. Elastin is the most flexible but the easiest to damage. The ground substance is just as important for determining the function of a tissue as the fibers are. There is much more variation here than in the fibers, and so the best approach is to examine each as we delve into the individual tissues.
7.3 Bone and Cartilage Components
There are many players in the building of a skeleton, but for the purposes of brevity, we will discuss only a few major different types: bone, hyaline cartilage, elastic cartilage, and fibrocartilage. Most of this discussion will focus on hyaline cartilage and bone, as they are the most abundant tissues found in the skeleton. They are key components of the adult skeleton and also play a major role in the development of the skeleton.
When considering the skeleton, bone is the first tissue that comes to mind. Bone is one of the strongest biological materials known. In compression, it can withstand over 20,000 psi. The majority of vertebrate groups use bone as the foundation of their skeleton.
Bone has four major cell types: osteoblasts, osteocytes, osteogenic cells, and osteoclasts (Figure 7.2). They are responsible for building, maintaining, and breaking down bone. The major fiber in bone is collagen. The ground substance in bone has two major components: an organic matrix and an inorganic matrix. The organic matrix, known as osteoid, helps connect the cells embedded in bone, the collagen fibers, and the inorganic matrix. The inorganic elements of the matrix are known as hydroxyapatite. The bulk of the hydroxyapatite is mineral salts, particularly those of calcium and phosphate. The mineral salts compose almost two-thirds of the total mass of bone. It is important to consider the calcium and phosphate together with the collagen fibers in understanding the functional strengths and limits of bone. The hydroxyapatite is extremely strong but extremely brittle. The collagen woven through the ground substance provides bone with some flexibility as well as increasing bone’s strength in tension. Without the collagen and the hydroxyapatite, the function of bone is greatly impaired. This is especially important in the context of human biology and medicine. As we age, the rate at which tissues are replaced slows down. The collagen in our bones loses its limited flexibility, and the bones become more brittle, increasing the likelihood that they will fracture. Additionally, changes in calcium availability and replacement begin to impact the strength of the bone. A lack of calcium in the diet, along with hormonal changes due to aging, can result in the loss of bone density in a condition we refer to as osteoporosis. Osteoporotic bones are much more likely to fracture when subjected to even minor impacts or forces.
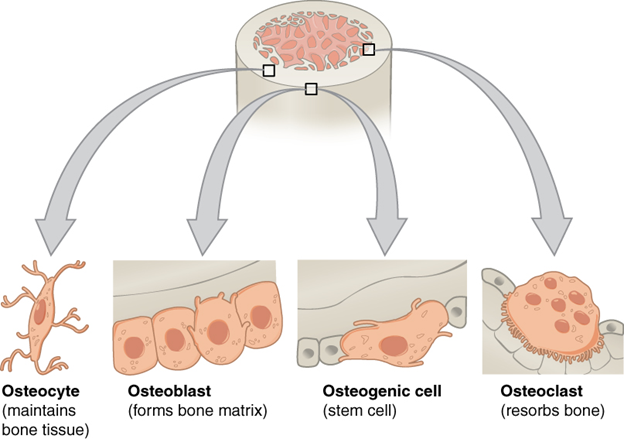
Figure 7.2—The major cell types of bone cells.
Hyaline cartilage is the most common cartilage in the body. Much like bone, it is a connective tissue, so we can identify the key cells and extracellular matrix that will define hyaline cartilage and how it functions (Figure 7.3).
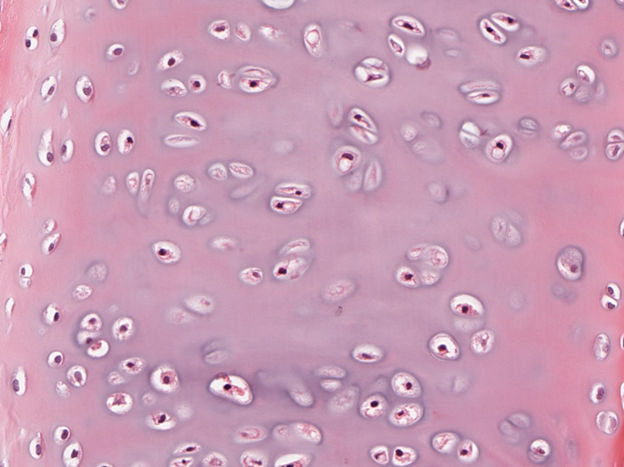
Figure 7.3—Hyaline cartilage of the trachea. Note the small chondrocytes surrounded by the purple/pink staining ECM.
All of the cartilages have two major types of cells, the chondroblasts and chondrocytes. Chondro- always refers to cartilage. The -blasts and -cytes work similarly to how they did in bone. Chondroblasts are immature cells that build cartilage. Chondrocytes are embedded within the extracellular matrix, monitoring the mature cartilage for damage. There are chondroclasts, cells that may break down and resorb cartilage, but they are rare. In fact, it’s only been within the last 10 years that we have started to pin down what chondroclasts are and where they originate. The extracellular matrix for hyaline cartilage is full of collagen fibers, with some elastin. This gives hyaline a little more flexibility than bone. The ground substance is also more flexible than bone. It is composed of three major types of molecules: glycosaminoglycans, proteoglycans, and glycoproteins. These sugar-protein combinations absorb water when placed in the extracellular matrix and help bind the collagen to the rest of the matrix. This results in a material that is tough (but not as strong as bone) but also fairly flexible. As a result of these properties, you can find hyaline cartilage in many places that require both toughness and flexibility. The ends of many bones have a layer of hyaline cartilage, particularly those involved in joints, which we refer to as articular cartilage. The costal cartilages, which connect the ribs to the sternum in many animals, are also hyaline cartilage. There is hyaline cartilage in nonskeletal locations as well; the cartilage that keeps the trachea open is hyaline. Later in this chapter, when we discuss the development of the skeleton, we will learn that many bones originate as hyaline cartilage before they are replaced with true bone. In humans, we can find remnants of this system in our growth plates, also known as epiphyseal plates, where a bit of hyaline cartilage remains and is the center for bone growth.
Elastic cartilage and fibrocartilage (Figure 7.4) play more reduced roles in the body than hyaline cartilage or bone. Elastic cartilage is not involved in the skeleton, found predominantly in the external ears and the tip of the nose in some mammals. It has the same general cell types as hyaline. The difference is in the fibers. Elastic cartilage is almost entirely elastic fibers, composed of a protein known as elastin (repetitive, isn’t it?). Elastin is the most flexible of all the fiber types, able to be deformed and stretched without damage. However, it is not the toughest material, and it is easily damaged. Fibrocartilage is more common, found in several places: the intervertebral discs between individual vertebrae, menisci in the knees of many vertebrates, and symphyses (the pubic symphysis in humans, for example). All three of these structures sit between bones. In the case of the intervertebral discs, the fibrocartilage surrounds fluid that allows for additional cushioning. Menisci sit between the tibia and femur of most tetrapods and also provide cushioning to the knee joint. Symphyses are fibrocartilage connections between bones. The pubic symphysis provides a small amount of flexibility between the left and right sides of the pelvis.
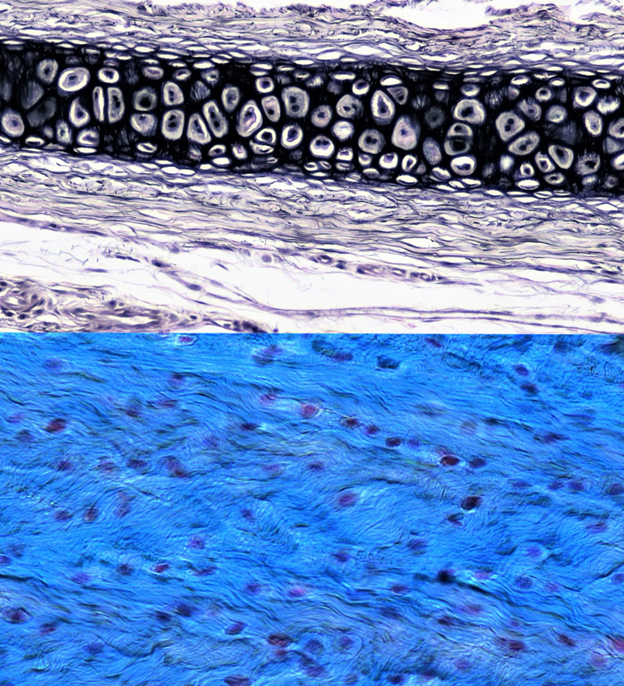
Figure 7.4—Top: Elastic cartilage. Bottom: Fibrocartilage.
7.4 Bone Structure
Now that we have the cellular components of the skeleton, we can look to individual bones for additional information on how the skeleton is constructed and how it functions. Individual bones can be broken down into several different regions. None of the bones are completely solid and instead may have channels or even large cavities within them. Using the human femur as an example (Figure 7.5), we can see the regions of a bone.

Figure 7.5—A human femur with major structures labeled.
The largest section of this bone, sometimes referred to as the shaft of the femur, is known as the diaphysis. The diaphysis has a large cavity at its center, often referred to as the marrow cavity. This is where red or yellow bone marrow can be stored, red for the formation of blood cells and yellow for the storage of fats and nutrients. At each end of the bone are the epiphyses. The epiphyses are important for bone development as well as bone growth. Within the epiphyses we find the previously mentioned epiphyseal plates, or growth plates as we know them in people. Before sexual maturation, the epiphyseal plates are primarily hyaline cartilage and responsible for the growth of bone in length. After maturity, the growth plates fuse and become entirely bone. There are exceptions to this, as many animals can continue to grow in size beyond sexual maturity. Beyond the growth plates, the epiphyses are another place to identify two types of bone that are found throughout the body. When viewed from its external surfaces, bone appears to be quite solid. This is actually a specific type of bone, known as compact bone. It is, in fact, quite solid but not continuous throughout the entire bone. In the diaphysis of the long bone, compact bone is found on the periosteal (external) and endosteal (internal) surfaces. In the epiphyses, it may only appear on the periosteal surface. The internal element is composed of a porous material full of channels and bone struts. This is trabecular bone (also referred to as spongy or cancellous bone). Trabecular bone provides the maximum amount of strength with the minimal amount of material. Trabecular bone is heavily regulated and constantly remodeled. The patterns of trabecular bone can be used to understand the forces encountered during movement in an area. The lines of the trabecular bone may match the lines of force.
7.5 Bone Development and Growth
There are two major patterns of bone development: endochondral and intramembranous. The development of the skeleton is a very well-understood process but a little beyond the scope of our work here. Instead, we can briefly discuss the two processes, where they occur, and who the major players are. Intramembranous ossification (making bone) is typically associated with flat bones like the skull and sternum (Figure 7.6).
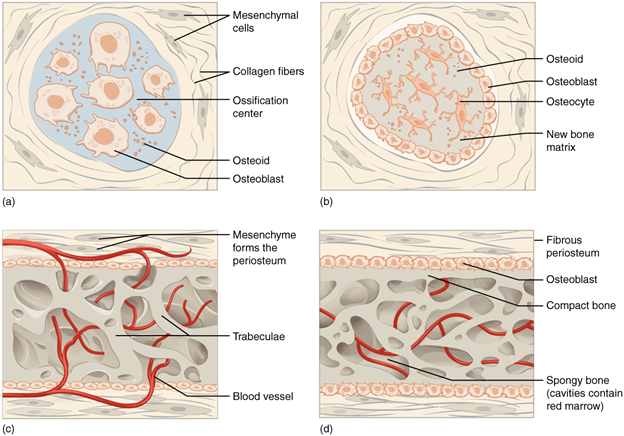
Figure 7.6—The process of intramembranous ossification. Stages occur in order from A to D.
Take a look at a human skull, and find the parietal bone, as it is formed this way. Early in development, mesenchymal cells find their way in between sheets of connective tissue (Figure 7.6A). Responding to environmental and genetic signals, they become osteoblasts and begin to secrete the osteoid we discussed earlier (Figure 7.6B). With many of these cells working, some get trapped in the bone as it is being laid down. These trapped osteoblasts mature into osteocytes and begin the work on making sure the bone is properly set up. As more and more bone is placed, osteoclasts move in to create openings for nerves and blood vessels (Figure 7.6C). The process continues until the bone is almost fully formed (Figure 7.6D). This pattern of development is primarily used for these flat bones; intricate shapes (like those of the femur/humerus) are not conducive to this type of development. For these bones, the process of endochondral ossification is more common. Endochondral roughly translates to “within cartilage,” and this is how the process unfolds. You can see the overall process in Figure 7.7. The long bones, like the humerus and femur, begin as cartilage molds (there’s that pesky hyaline again; Figure 7.7A). Slowly, that cartilage is replaced by bone.
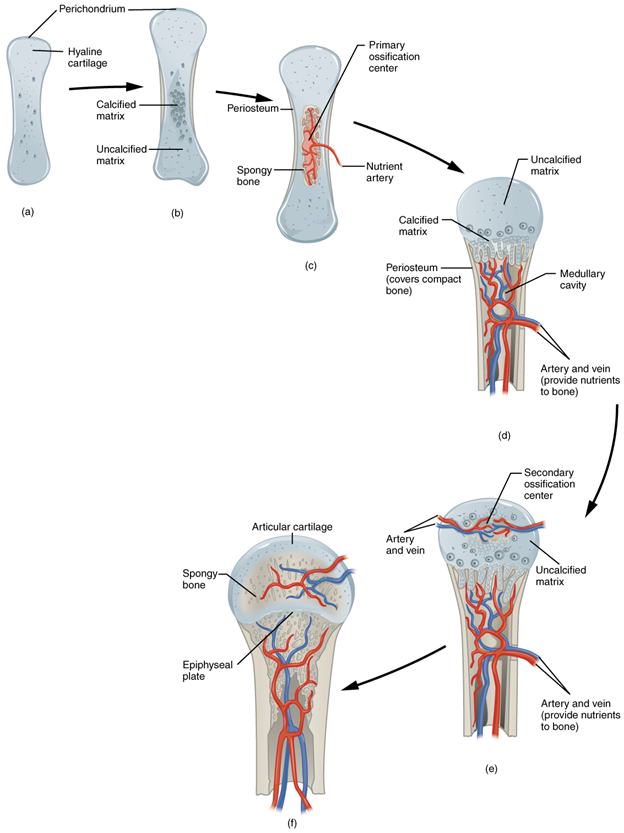
Figure 7.7—The process of endochondral ossification in a human long bone.
The hyaline cartilage is calcified (Figure 7.7B), and as the chondroblasts and chondrocytes die, they are replaced by osteoblasts. Note the differences in location of the primary and secondary ossification centers as well as their fates (Figures 7.7C and 7.7E). The primary ossification center will be the diaphysis, with the marrow cavity as its center (Figure 7.7D). The secondary ossification centers will become the epiphyses. As these areas grow, they converge toward one another. However, they don’t quite meet. Before birth, these centers stop growing toward one another, leaving a small area of cartilage between them (Figure 7.7F). If this sounds familiar, that is because we recently discussed these areas. These are the epiphyseal growth plates!
Now that the bones have formed, it is easy to think that they will remain the same throughout life. In fact, this couldn’t be further from the truth. As an individual animal grows, the bones must grow with them. We can consider bone growth in two different processes: interstitial growth (growth in length) and appositional growth (growth in width). Interstitial growth occurs at the epiphyseal growth plates. At the center of those growth plates, hyaline cartilage remained. The chondroblasts and chondrocytes are still active, building new cartilage. As they do so, they push the epiphysis and diaphysis farther from each other, making the bone incrementally longer. As the cartilage moves farther from the center, it starts to become ossified. Calcium and phosphorus salts become embedded in the ECM, and the chondroblasts and chondrocytes die. When they do, this makes space for osteoblasts and osteocytes to move in and convert the ossified cartilage into new bone. Simultaneously, the osteoclasts will be removing excess bone as it moves farther from the epiphyseal growth plate. Appositional growth is much simpler. On the periosteal (external) surface of the bone, osteoblasts are adding new bone, while osteoclasts work on the endosteal surface to remove bone. This makes the bone wider, and as long as the osteoblasts are laying down at a higher rate than the osteoclasts can remove it, the bone will also get relatively thicker. This is particularly important for the large weight-bearing bones, like the femur and tibia.
After sexual maturity, many (but not all) vertebrates stop growing in length/height. The epiphyseal plates fully ossify into bone (in humans, this is in response to the spike in sex hormones), “fusing” the epiphyses and diaphyses together. However, that does not mean that the skeleton is done modifying for the rest of an animal’s life. Bone is an extremely dynamic tissue, constantly being turned over, replaced, and changed in response to the needs of an organism. We call this process remodeling, much in the way you might remodel your bedroom or a kitchen. It is easiest to consider remodeling in the context of humans. Earlier, we described the spongy bone, or trabeculae, as allowing for maximal stress while minimizing material costs and weight. The trabeculae are laid down along lines of stress, dependent on how the bone is being used. Try this exercise: Find a partner, and have them jog in place. Notice how vertical the femur (their large thigh bone) remains while they are jogging. Now have them squat as low as they can. See how the femur moves into a more horizontal position. For these two activities, the body weight of the person doing the movement travels to the ground along different “lines.” If they were a marathon runner or weight lifter, you can imagine that the stresses of their activities would result in bones that look very different. These would be reflected in their trabeculae as well as the thickness of the compact/cortical bone. However, if the marathon runner gives up their sport, for whatever reason, over time the bones will remodel to their new activity pattern. This is a matter of concern for many older adults who struggle to maintain active lifestyles. A lack of activity or becoming more sedentary will often result in a net loss of bone, as the bone that was necessary for movements or stresses is no longer needed and thus removed. It can be restored, but care must be taken, as the process of rebuilding bone slows down significantly as we age.
7.6 Further Reading
- Sorenson, Robert L., and T. Clark Belje. Atlas of Human Histology: A Guide to Microscopic Structure of Cells, Tissues and Organs. Minneapolis: University of Minnesota Press, 2014.
- “Cartilage and Bone.” Histology Guide, n.d. https://histologyguide.com/slidebox/05-cartilage-and-bone.html.
7.7 References
- Betts, J. Gordon, Kelly A. Young, James A. Wise, Eddie Johnson, Brandon Poe, Dean H. Kruse, Oksana Korol, Jody E. Johnson, Mark Womble, and Peter DeSaix. Anatomy and Physiology 2e. 2024.
- Caetano-Lopes, Joana, Helena Canhao, and Joao Eurico Fonseca. “Osteoblasts and bone formation.” Acta Reumatológica Portuguesa 32, no. 2 (2007): 103–110.
- Dean, Mason N., and Adam P. Summers. “Mineralized cartilage in the skeleton of chondrichthyan fishes.” Zoology 109, no. 2 (2006): 164–168.
- DeSaix. “4.3 Connective Tissue Supports and Protects—Anatomy and Physiology 2e.” OpenStax, April 20, 2022. https://openstax.org/books/anatomy-and-physiology-2e/pages/4-3-connective-tissue-supports-and-protects.
- DeSaix. “6.3 Bone Structure—Anatomy and Physiology 2e.” OpenStax, April 20, 2022. https://openstax.org/books/anatomy-and-physiology-2e/pages/6-3-bone-structure.
- DeSaix. “6.4 Bone Formation and Development—Anatomy and Physiology 2e.” OpenStax, April 20, 2022. https://openstax.org/books/anatomy-and-physiology-2e/pages/6-4-bone-formation-and-development.
- Iqbal, Jameel, and Mone Zaidi. “Endothelial cells revealed as chondroclasts.” Nature Cell Biology 21, no. 4 (2019): 417–419.
- Lewinson, Dina, and Michael Silbermann. “Chondroclasts and endothelial cells collaborate in the process of cartilage resorption.” Anatomical Record 233, no. 4 (1992): 504–514.
- Mackie, E. J., Y. A. Ahmed, L. Tatarczuch, K-S. Chen, and M. Mirams. “Endochondral ossification: How cartilage is converted into bone in the developing skeleton.” International Journal of Biochemistry & Cell Biology 40, no. 1 (2008): 46–62.
- Mizoguchi, Toshihide, and Noriaki Ono. “The diverse origin of bone-forming osteoblasts.” Journal of Bone and Mineral Research 36, no. 8 (2020): 1432–1447.
- Oláh, Tamás, Tunku Kamarul, Henning Madry, and Malliga Raman Murali. “The illustrative anatomy and the histology of the healthy hyaline cartilage.” The Illustrative Book of Cartilage Repair, edited by Deepak Rajkumar Goyal, 5–10. New York: Springer, 2021.
- White, Alicia, and Gillian Wallis. “Endochondral ossification: A delicate balance between growth and mineralisation.” Current Biology 11, no. 15 (2001): R589–R591.
